Developmental hematopoietic stem cell fate decisions explain clonal dynamics over lifetime
Behind the Paper
Developmental hematopoietic stem cell variation explains clonal hematopoiesis later in life
Jesse Kreger, Jazlyn A. Mooney, Darryl Shibata, Adam L. MacLean
Read the paperClonal expansion of blood stem cells (clonal hematopoiesis or CH) is a hallmark of aging. Blood cancers such as leukemia or myelodysplastic syndrome can develop from such clonal expansions. However, CH without any clinical implications is observed in a large fraction of the aging population. Why is CH pervasive in healthy individuals? What are the origins of CH in the blood? These questions – raised by several recent studies – are of importance not only to understand the clonal composition of the blood and associated disease risks, but also shed light on the evolution of somatic variants throughout a human life.
Here, we provide an answer to the question of the origins of CH[1]. We show that stem cell variants arising during development are sufficient to explain the emergence of CH many decades later as we grow old. Although driver mutations are present in some blood diseases of clonal origin, we show how CH can be pervasive in many cases lacking any known driver mutations.
This paucity of driver mutations in most cases of CH has been shown recently via several deep sequencing studies[2,3]. Serial observations of hematopoiesis are consistent with weak selection because clone sizes are often stable or expand slowly. However, weak selection seems implausible to explain CH since stem cell variants that arise in the bone marrow with weak selective advantages would likely be randomly lost due to stochastic cell replacement. Yet: we observe the widespread emergence of CH in individuals over the age of 65.
In this work we propose a simple theory that resolves this paradox, and which is fully consistent with the data: stem cell variation leading to CH in later life is present before birth. We provide unique means with which to test this theory via two essential components. First, whereas deep DNA sequencing leads to insight into the mutational history of cells throughout life, it cannot resolve variation arising in development due to the relatively few mutations occurring at this time. In contrast, higher rates of fluctuating methylation during development allow stem cells to become polymorphic by birth and – as has been previously shown – enable lineage tracing throughout life[4]. Second, twin studies allow us to quantify differences in clonal distributions that occur due to genetic similarity or shared blood circulation in utero, in the case of some monozygotic twin pairs (Figure 1).
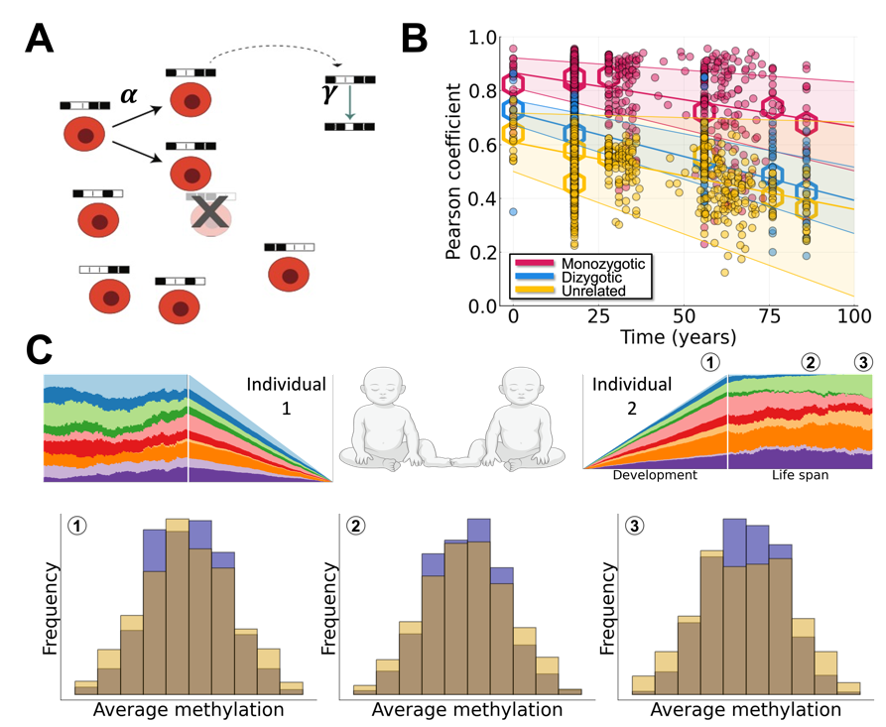
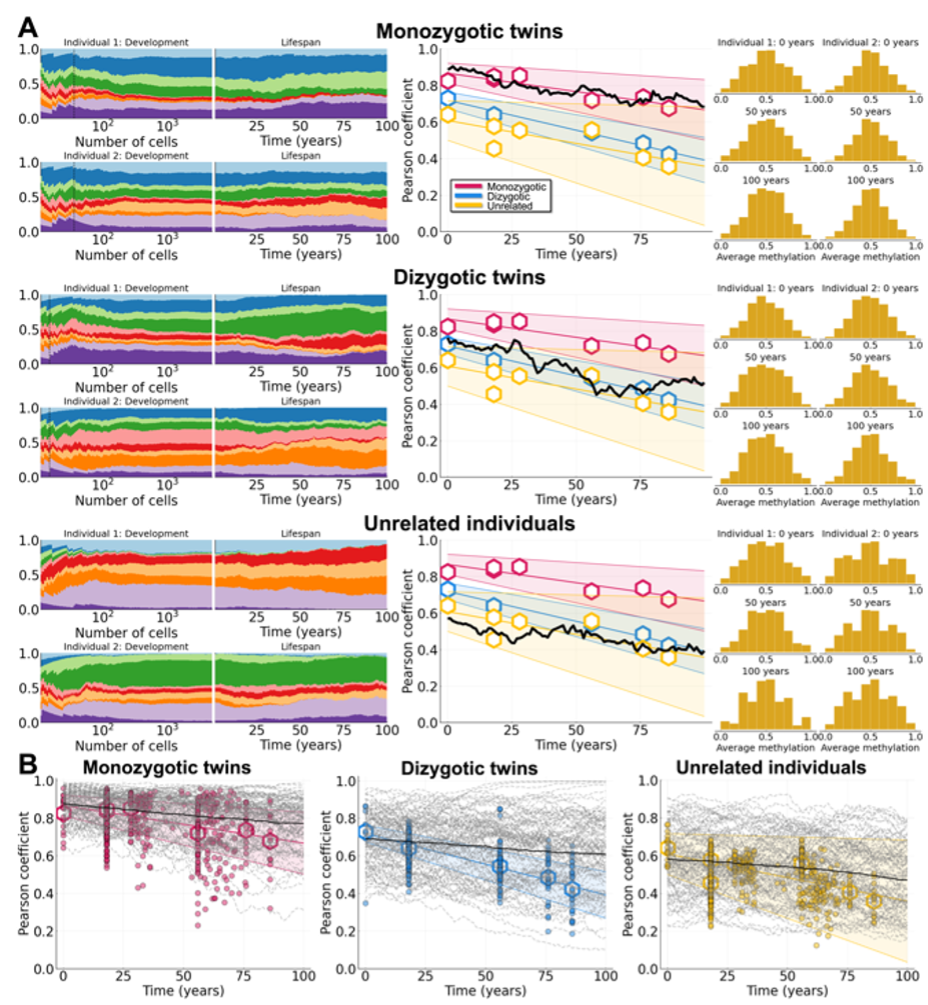
To test this theory, we developed a mathematical model of hematopoietic stem cell somatic evolution that describes clonal dynamics in development and throughout life. Cell replacement is modeled with a Moran model and fluctuating CpG sites are described by a Markov process. Using the fluctuating methylation dynamics as a lineage marker, we fit the model to data describing the dynamics of twins vs unrelated individuals, using a large compendium of datasets encompassing clonal information from birth until old age (~82 years old). Following parameterization of the model to data, we show that variants that arise before birth conferring weak selective survival advantages commonly become dominant later in life (Figure 2). I.e. developmental origins of blood stem cell variation reconciles the controversy raised by the lack of driver mutations found in many recent studies, and demonstrates that many blood subclones that grow to large sizes later in life were likely created before birth.
Of course, there remains much to be done. Stem cell aging in the bone marrow involves many different factors. Here, we have investigated the clonal origins of stem cell heterogeneity; specific genetic mutations[5], as well as environmental[6] (i.e cell-extrinsic), epigenetic[7], transcriptional and metabolic[8] changes also play crucial roles in stem cell aging. Future models ought to begin the challenging yet important task of integrating – perhaps not all but – several of these factors into consistent models that are able to describe and predict phenotypes of aging in the blood. While this may not lead to a reversal of aging (as some billionaires aspire[9]), we hope it leads to new understanding and tools with which to extend quality of life by maintaining a balanced and well-functioning population of stem cells in the bone marrow.
References
- Kreger, J. et al. Developmental hematopoietic stem cell variation explains clonal hematopoiesis later in life. bioRxiv (2024). https://www.biorxiv.org/content/10.1101/2024.03.02.583106
- Stacey, S.N. et al. Genetics and epidemiology of mutational barcode-defined clonal hematopoiesis. Nature Genetics (2023). https://www.nature.com/articles/s41588-023-01555-z
- Mitchell, E. et al. Clonal dynamics of haematopoiesis across the human lifespan. Nature 606, 343–350 (2022). https://www.nature.com/articles/s41586-022-04786-y
- Gabbutt, C. et al. Fluctuating methylation clocks for cell lineage tracing at high temporal resolution in human tissues. Nature Biotechnology 40, 720–730 (2022). https://www.nature.com/articles/s41587-021-01109-w
- Schenck, R.O. et al. Homeostasis limits keratinocyte evolution. PNAS 119, e2006487119 (2022). https://www.pnas.org/doi/full/10.1073/pnas.2006487119
- Young, K. et al. Decline in IGF1 in the bone marrow microenvironment initiates hematopoietic stem cell aging. Cell Stem Cell 28, 1473-1482 (2021). https://www.cell.com/cell-stem-cell/abstract/S1934-5909(21)00123-5
- Rommelfanger, M.K. et al. Gene regulatory network inference with popInfer reveals dynamic regulation of hematopoietic stem cell quiescence upon diet restriction and aging. bioRxiv (2023). https://biorxiv.org/content/10.1101/2023.04.18.537360
- Tang, D. et al. Dietary restriction improves repopulation but impairs lymphoid differentiation capacity of hematopoietic stem cells in early aging. J Exp Med 213, 535-553 (2016). https://rupress.org/jem/article/213/4/535/41962/Dietary-restriction-improves-repopulation-but
- Klee, M. Millionaire Biohacker Says Algorithm Runs His Life: ‘My Mind No Longer Decides’ (2023, September 11). Rolling Stone. https://www.rollingstone.com/culture/culture-features/bryan-johnson-anti-aging-blueprint-algorithm-1234821163. [Accessed: 2024, April 9]
© 2026 - The Mathematical Oncology Blog