Lenia: Bridging Artificial Life and Cancer Biology
Behind the paper
Spatial interactions modulate tumor growth and immune infiltration
Sadegh Marzban, Sonal Srivastava, Sharon Kartika, Rafael Bravo, Rachel Safriel, Aidan Zarski, Alexander R. A. Anderson, Christine H. Chung, Antonio L. Amelio & Jeffrey West
Read the paperLenia & artificial life
How cool is the name 'Artificial Life'? It literally means bringing life to a computer, letting you test your craziest ideas without any of the real-world side effects! We used one such system, Lenia[1], to explore tumor growth and study interactions within the tumor-immune environment. Lenia (from the Latin lenis, meaning “smooth”) is a cellular automata framework that operates with continuous space and time, inspired by Lenia Mathematical Lifeforms—a continuous generalization of John Conway’s Game of Life. Before going into the technical details, it’s important to reflect on why we created this model. Artificial life systems are designed to capture essential biological features such as self-organization (morphogenesis), self-regulation (homeostasis), self-direction (motility), and self-replication (reproduction). They also model processes like growth, response to environmental stimuli, evolvability, and adaptation—emergent properties critical in living systems. These same characteristics—cellular replication, growth, motility, evolvability, and adaptation—are not only hallmarks of biological life but also key drivers in the development of complex, multi-factorial diseases such as cancer. By modeling these processes, Lenia allows us to simulate the intricate behaviors of cancer cells, providing deeper understanding into how tumors grow, evolve, and respond to treatment. Let’s explore some examples using Lenia.Logistic Growth with an Allee effect
In our recent paper[2], we first imagine a small group of tumor cells in the early stages of growth. At this point, there may not be enough cells to release sufficient growth factors, evade the immune system, or establish a proper blood supply (angiogenesis). As a result, the tumor grows slowly, or it might even shrink. However, once the tumor reaches a critical size, there are enough cells to start processes like angiogenesis, which brings nutrients to the tumor. This increased communication between cells helps the tumor grow more rapidly. Thus, there’s a threshold, called the Allee threshold, that, once crossed, speeds up tumor growth. In tumor growth dynamics literature, a logistic growth model (Equation 1) is often used. This model creates an "S-curve," where the tumor initially grows like it would under exponential growth when it's far from its maximum size (carrying capacity). But as it gets closer to the carrying capacity, growth slows down due to resource limitations. $$ \begin{equation} \frac{dN}{dt}=γN(1- \frac{N}{k}) \\ \end{equation} $$ To account for the possibility that tumors might go extinct early on—due to the Allee effect—we can introduce the Allee threshold into the model (Equation 2). Figure 1 compares how tumor growth behaves under the basic logistic model, a small Allee threshold, and a large one. Figures 1(A-C) show that having a higher initial number of tumor cells can help avoid early extinction. $$ \begin{equation} \frac{dN}{dt}=γN(1- \frac{N}{k})\frac{(N-L)}{k} \end{equation} $$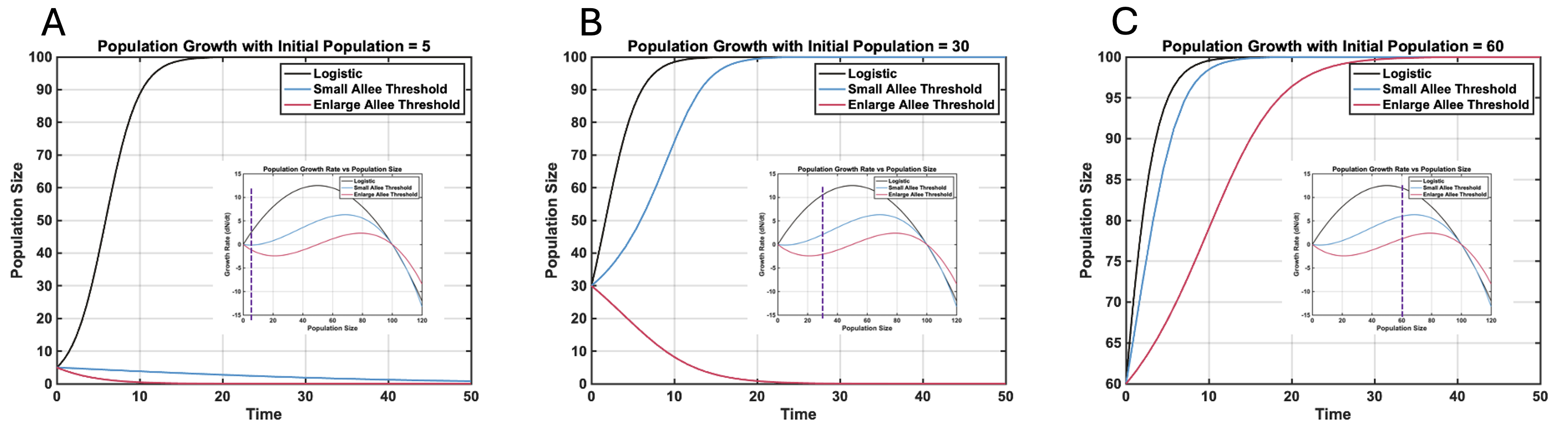
“Short-range interaction kernels are more robust to Allee effects.”These models don’t fully capture the heterogeneity and interactions within the tumor microenvironment. For instance, when considering the Allee effect, we might want to explore how this threshold interacts with spatial dynamics in tumors. More precisely, the interaction kernel of a tumor accounts for a spectrum of spatial interactions, ranging from local (cell-scale) to global (tumor-scale). We’re interested in understanding how this range of interactions can affect tumor dynamics across different Allee thresholds.
Spatial Interactions and Heterogeneity in Tumor-Immune Dynamics
While the logistic and Allee models provide a foundation for understanding tumor growth, they don’t fully capture the spatial heterogeneity within the tumor microenvironment. Lenia offers a unique advantage by supporting multi-interaction kernels in the system. In the context of tumor-immune interactions, different types of interactions—such as tumor-tumor, tumor-immune, immune-tumor, and immune-immune—can each influence the dynamics in different ways. These interaction kernels represent the spatial dynamics and distances over which cells interact. To describe tumor-immune dynamics in this more complex setting, we turn to a predator-prey model described in Equations (3) and (4). Tumor cell growth dynamics: $$ \begin{equation} \dot{x}=rx(x-L)(1-x)-bxy \end{equation} $$ Immune cell dynamics: $$ \begin{equation} \dot{y}=gbxy-dy \end{equation} $$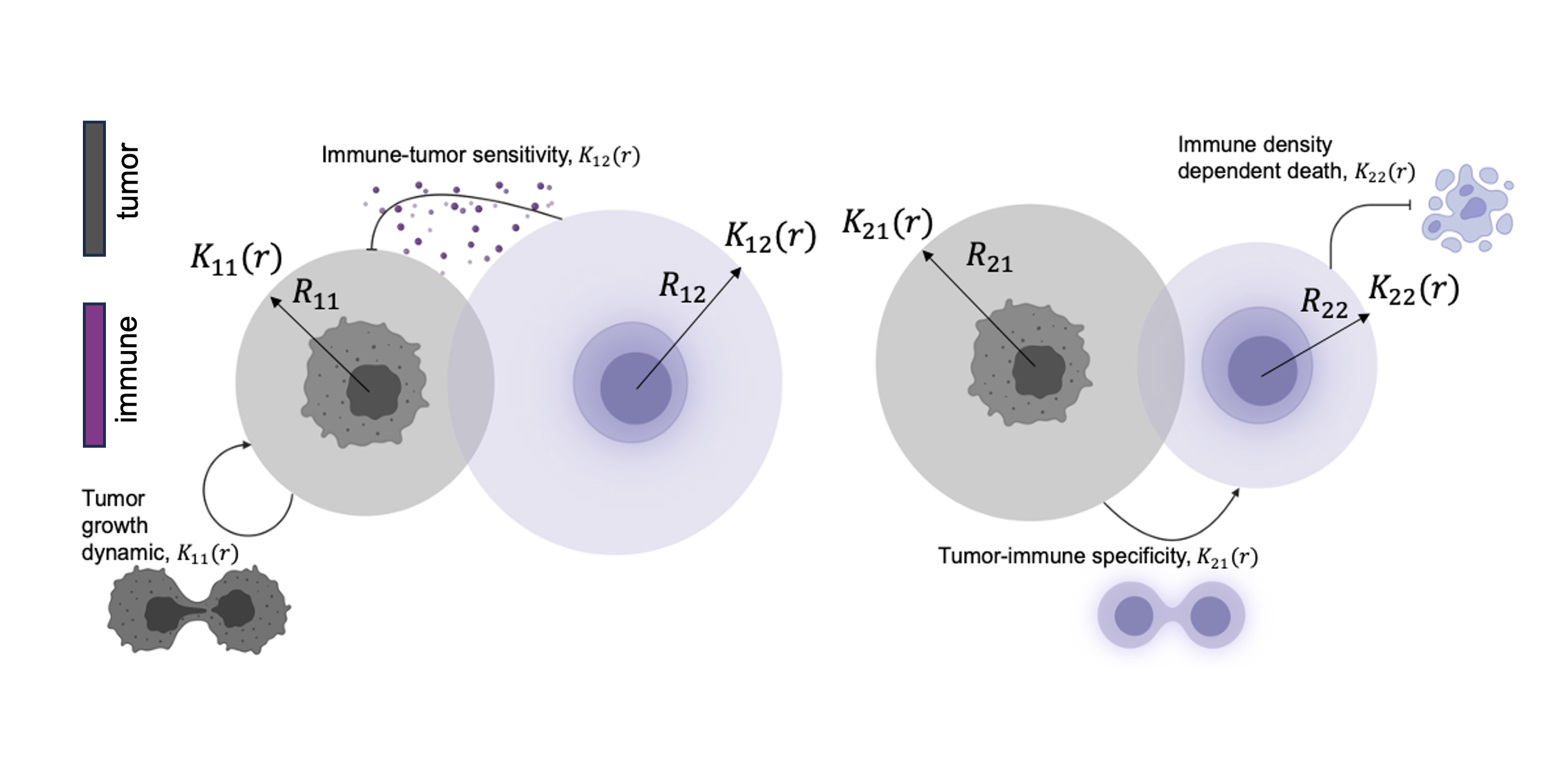
- Classical Analytical Models: Simulating ordinary differential equations widely applied in cancer research.
- Stochastic Agent-Based Models: Supporting stochastic simulations of cancer cell behavior.
- Evolutionary Game Theory: Modeling interactions between multiple cell types competing on Darwinian fitness landscapes.
- Cell Migration: Incorporating generalized cell migration models like chemotaxis, which are prevalent in mathematical modeling of cancer.
Mini-tutorial for Lenia
If you'd like to use Lenia in your project or recreate the simulations shown in this blog, you can easily download and implement it via github.com/mathonco/Lenia-in-hal, built in HAL[3]. Users can define growth functions and customize kernel sizes and interaction functions for spatial modeling. Implementing a model in Lenia involves two key steps:- Defining the growth function, (G(u))
- Defining the interaction kernel, (K(r))
// Growth function
@Override
public double G(double u) {
return gamma * u * (u - L) * (C - u);
}@Override
public double K(double r) {
if (r <= Rstar) {
return 1.0;
} else {
return 0.0;
}
}References
- Chan, B.W.C., 2018. Lenia-biology of artificial life. arXiv preprint arXiv:1812.05433.
- Marzban, S., Srivastava, S., Kartika, S., Bravo, R., Safriel, R., Zarski, A., Anderson, A.R., Chung, C.H., Amelio, A.L. and West, J., 2024. Spatial interactions modulate tumor growth and immune infiltration. NPJ Systems Biology and Applications, 10(1), p.106.
- Bravo, R.R., Baratchart, E., West, J., Schenck, R.O., Miller, A.K., Gallaher, J., Gatenbee, C.D., Basanta, D., Robertson-Tessi, M. and Anderson, A.R., 2020. Hybrid Automata Library: A flexible platform for hybrid modeling with real-time visualization. PLoS computational biology, 16(3), p.e1007635.
© 2026 - The Mathematical Oncology Blog