The impact of phenotypic heterogeneity of tumour cells on treatment and relapse dynamics
Michael Raatz, Saumil Shah, Guranda Chitadze, Monika Brüggemann, Arne Traulsen
Read the preprint
It is often argued that mathematically motivated treatment should leverage ecological principals to overcome current treatment limitations, for example to avoid resistance evolution
1,5,6,9. One of the cornerstones of contemporary ecological research is the investigation of intra-specific heterogeneity and its consequences for populations and ecosystems
3,4. This stresses that individuals from the same species, even those that are closely related, can be different from each other in many aspects. Sometimes, individuals from different species can be more similar than individuals from the same species.
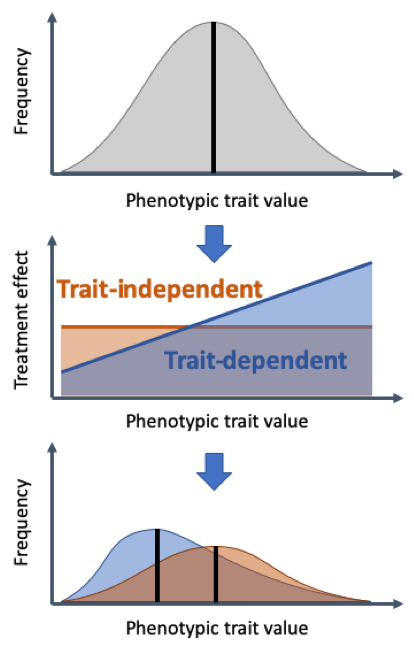
Characterizing this heterogeneity is achieved by measuring functional traits of organisms, thus obtaining trait distributions for populations in a research branch called trait-based ecology. Notably, these trait distributions are not necessarily binary, but allow to cover broad spectra of differences between organisms. The term functional is here synonymous to “having an effect” and describes that a focus is put on heterogeneity in those traits that can mechanistically be attributed to certain effects. As origins of this heterogeneity not only evolutionary changes, but also phenotypic plasticity are considered. For example, both evolution and phenotypic plasticity can drive the deployment of anti-predator defenses in prey.
While intratumour heterogeneity is more and more recognized as biological and medical reality with prognostic value
7, using trait-based models in cancer modelling is rather uncommon. In particular, non-binary descriptions of heterogeneity are rare. In our recent
preprint11, we implemented a trait-based description of a heterogeneous cancer cell population and investigated how it responds to different treatment types. We deployed both a deterministic description with ordinary differential equations to further our mechanistic understanding as well as a stochastic model to capture the randomness involved in treatment dynamics. In our model, we consider growth rate as the focal trait that captures most of the heterogeneity between cells. We split the cancer cell population into subpopulations of different growth rates and include switching to adjacent subpopulations proportional to the growth rate. Having set up the model, we investigate two different types of treatment that either do or do not select on the trait heterogeneity. The cancer cell mortality exerted by the growth-rate-dependent treatment increases for faster-growing subpopulations, while is it constant for the growth-rate-independent treatment.
We find that the growth-rate-dependent treatment results in a smaller probability of tumour eradication, but if treatment fails the relapse is delayed and time to progression is larger. This apparent trade-off chimes with the current discussion about whether to aim for cure or for tumour containment
8,10. In our model, the mechanism behind this dichotomy lies in the dynamics of the trait distribution. The growth-rate-dependent treatment selects for slower growing subpopulations that dominate the cancer cell population towards the end of treatment. Due to their decreased treatment-induced mortality, such slow growing cells are more likely to survive the treatment phase, thus lowering the probability to eradicate all cancer cells in the growth-rate-dependent treatment compared to the alternative, growth-rate-independent treatment. At the same time, the fast-growing cells experienced a much higher mortality, reducing their subpopulations to much smaller numbers, driving many of them extinct. To regrow at high speeds, these faster subpopulations first have to be repopulated from the slower-growing subpopulations, which prolongs the time to progression compared to the growth-rate-independent treatment where the fast-growing subpopulations remain much higher at the end of treatment.
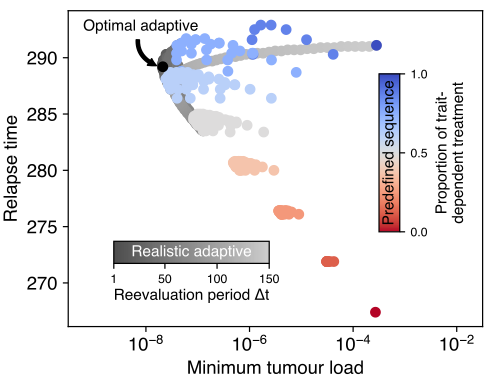
With the two different treatment types at hand, we next set off to investigate how their respective strengths may be leveraged. The growth-rate-dependent treatment is most effective when the trait distribution is skewed towards higher growth rates. Applying it, however, reduces the mean of the trait-distribution, which decreases the induced cell mortality, but eventually drives the population to territory where the growth-rate-independent treatment is superior. Switching the treatment type at that point thus ensures continued high mortality, even though now the (unselected) trait distribution will move toward faster growth rates again, motivating a reverse treatment change. Accordingly, we find that continued switching between both treatment types minimizes the tumour load at the end of treatment (thus maximizing the probability of cure) and ranks high with respect to delaying relapse.
In line with others
1,11,12, we thus conclude that awareness of cancer cell heterogeneity and the effect of different treatment types on this heterogeneity allows to navigate potential trade-offs between wanted and unwanted treatment responses. Using trait distributions to represent cancer cell heterogeneity allows to capture the often non-binary switches between sensitivity and resistance to a particular treatment.
References
- Gatenby, R. A., Silva, A. S., Gillies, R. J., & Frieden, B. R. (2009). Adaptive Therapy. Cancer Research, 69(11), 4894–4903. https://doi.org/10.1158/0008-5472.CAN-08-3658
- Gatenby, R. A., Brown, J., & Vincent, T. (2009). Lessons from applied ecology: Cancer control using an evolutionary double bind. Cancer Research, 69(19), 7499–7502. https://doi.org/10.1158/0008-5472.CAN-09-1354
- Bolnick, D. I., Amarasekare, P., Araújo, M. S., Bürger, R., Levine, J. M., Novak, M., … Vasseur, D. A. (2011). Why intraspecific trait variation matters in community ecology. Trends in Ecology and Evolution, 26(4), 183–192. https://doi.org/10.1016/j.tree.2011.01.009
- Violle, C., Enquist, B. J., McGill, B. J., Jiang, L., Albert, C. H., Hulshof, C., … Messier, J. (2012). The return of the variance: Intraspecific variability in community ecology. Trends in Ecology and Evolution, 27(4), 244–252. https://doi.org/10.1016/j.tree.2011.11.014
- Basanta, D., & Anderson, A. R. A. (2013). Exploiting ecological principles to better understand cancer progression and treatment. Interface Focus, 3(4), 20130020. https://doi.org/10.1098/rsfs.2013.0020
- Korolev, K. S., Xavier, J. B., & Gore, J. (2014). Turning ecology and evolution against cancer. Nature Reviews Cancer, 14(5), 371–380. https://doi.org/10.1038/nrc3712
- Gay, L., Baker, A.-M., & Graham, T. A. (2016). Tumour Cell Heterogeneity. F1000Research, 5, 238. https://doi.org/10.12688/f1000research.7210.1
- Hansen, E., & Read, A. F. (2020). Cancer therapy: Attempt cure or manage drug resistance? Evolutionary Applications, 13(7), 1660–1672. https://doi.org/10.1111/eva.12994
- Burmeister, A. R., Hansen, E., Cunningham, J. J., Rego, E. H., Turner, P. E., Weitz, J. S., & Hochberg, M. E. (2020). Fighting microbial pathogens by integrating host ecosystem interactions and evolution. BioEssays, 1–13. https://doi.org/10.1002/bies.202000272
- Viossat, Y., & Noble, R. (2020). The logic of containing tumors. bioRxiv. https://doi.org/10.1101/2020.01.22.915355
- Raatz, M., Shah, S., Chitadze, G., Brüggemann, M., & Traulsen, A. (2020). The impact of phenotypic heterogeneity of tumour cells on treatment and relapse dynamics. BioRxiv.
- Acar, A., Nichol, D., Fernandez-Mateos, J., Cresswell, G. D., Barozzi, I., Hong, S. P., … Sottoriva, A. (2020). Exploiting evolutionary steering to induce collateral drug sensitivity in cancer. Nature Communications, 11(1), 1923. https://doi.org/10.1038/s41467-020-15596-z