Short-Range Migration Can Alter Evolutionary Dynamics in Solid Tumors
Behind the paper
Short-Range Migration Can Alter Evolutionary Dynamics in Solid Tumors
Youness Azimzade, Abbas Ali Saberi
Read the manuscript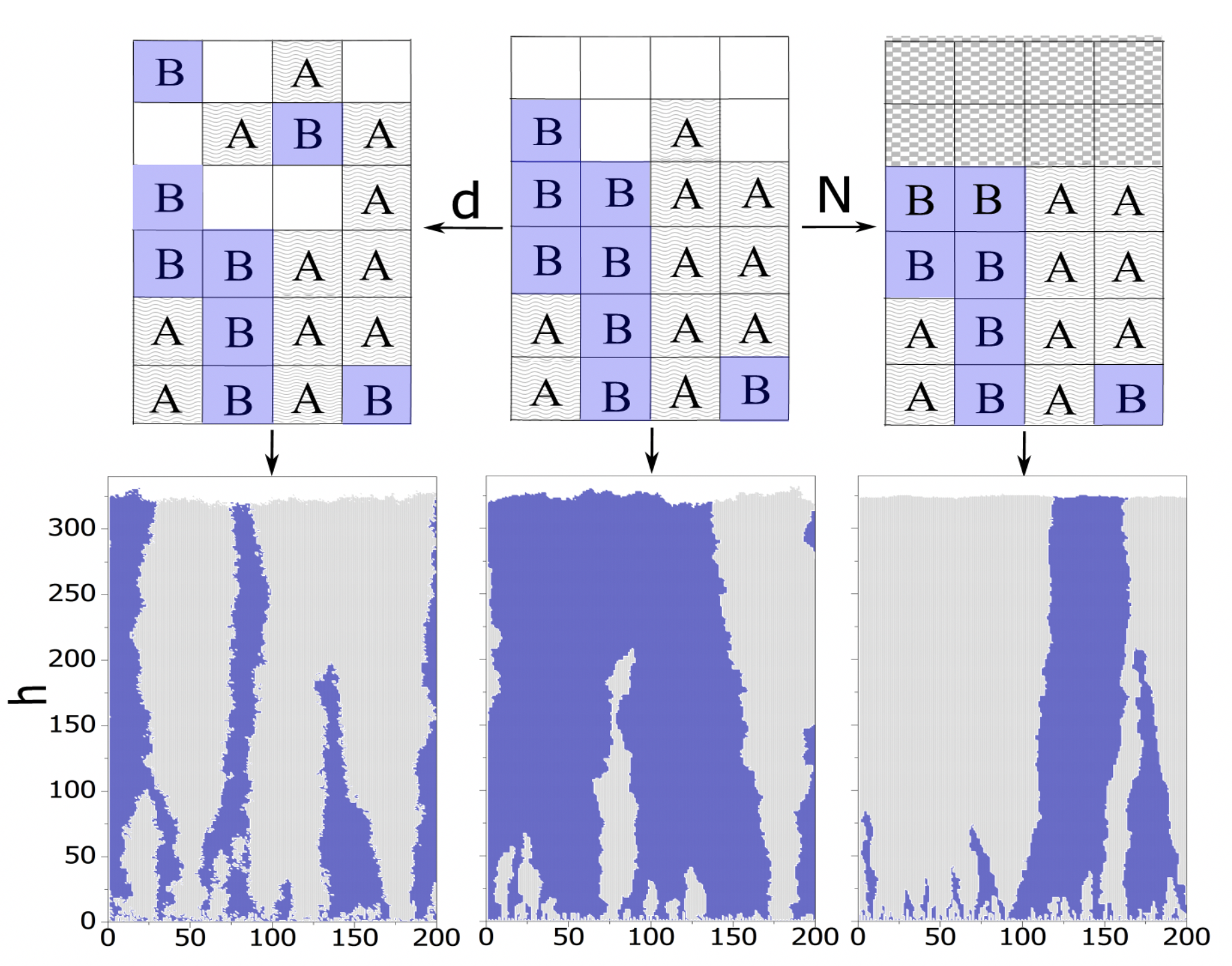 Our model prediction about the role of short-range migration is in line with recent findings on the effect of physical parameters on natural selection in bacterial colonies. Based on one of these studies, the collective migration of cells conceals fitness differences. However, our result predicts that even non-collective migration can increase the competition chance for the population with smaller fitness. FIG 2 depicts how such migration changes the efficiency of natural selection. In standard Eden model with R_B=1.1 R_A, (B) finally takes over the entire invasion front (FIG 2(a)). Adding removable physical barrier accelerates the fixation of (B) (FIG 2 (b)) and adding short-range migration weakens natural selection (FIG 2(c)).
Figure 2: Natural selection in presence of short range migration
Our model prediction about the role of short-range migration is in line with recent findings on the effect of physical parameters on natural selection in bacterial colonies. Based on one of these studies, the collective migration of cells conceals fitness differences. However, our result predicts that even non-collective migration can increase the competition chance for the population with smaller fitness. FIG 2 depicts how such migration changes the efficiency of natural selection. In standard Eden model with R_B=1.1 R_A, (B) finally takes over the entire invasion front (FIG 2(a)). Adding removable physical barrier accelerates the fixation of (B) (FIG 2 (b)) and adding short-range migration weakens natural selection (FIG 2(c)).
Figure 2: Natural selection in presence of short range migration
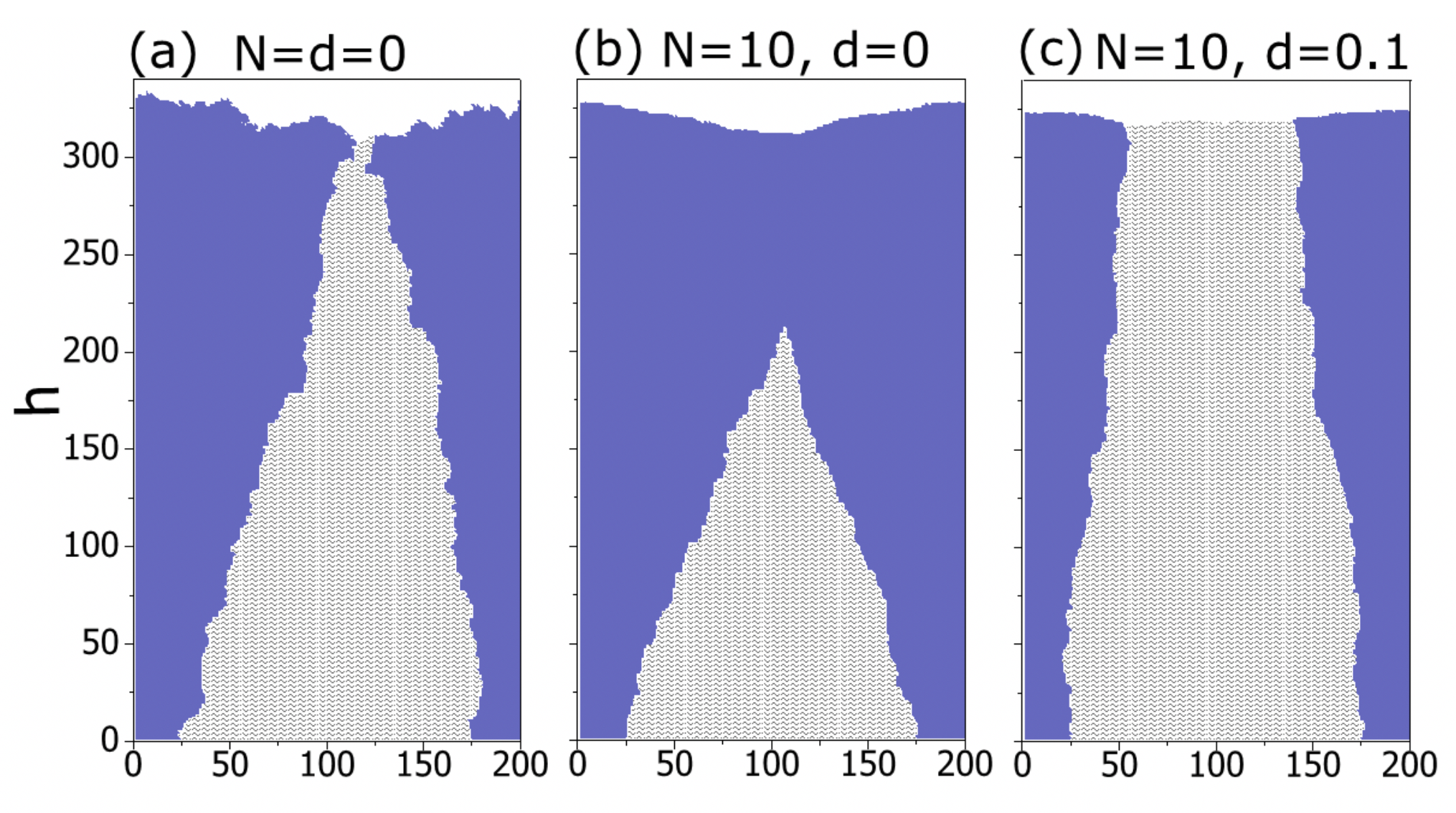 Our findings can be applied to different cellular aggregates such as bacterial colonies and tumors. However, we use this model to address the evolutionary dynamics in solid tumors. To show our findings in the context of tumor evolution, we use our 3D model to imitate tumor growth. We consider 3000 cells of (A) and (B) randomly positioned in the center of the 3D medium and let them grow according to the aforementioned rules. For two similar populations, both of them will appear at the surface and probable fixation is unlikely to happen due to the geometry of the tumor ( see FIG 3(a)). Then we consider the non-identical scenario with R_B=1.1 R_A. Based on the neutral theory, (A) should actively participate in the growth and should be present at the border of the tumor, in spite of having lower fitness. Based on Darwinian evolution, (B) is dominant and it will take over the whole tumor and will be the only population active in the border.
For the Eden model, as FIG 3(b) shows, (A) has a chance to appear at the border but finally it will be captured inside the tumor as the tumor becomes larger. When we add environmental barrier, the situation for (A) becomes worse and it will be run out of border faster (FIG 3(c)). However, when we add short-range migration by keeping N=10, the situation becomes completely different. First, two populations, as FIG 3(d) shows, became well-mixed. Then, in spite of being mixed, natural selection is not effective as before and (A) remains active in the border and participates in tumor evolution. Interestingly, while the dynamics of sub-clones has changed, the morphology of the tumor remains almost the same.
Figure 3: Evolutionary Dynamics of Tumor in presence of short-range migration
Our findings can be applied to different cellular aggregates such as bacterial colonies and tumors. However, we use this model to address the evolutionary dynamics in solid tumors. To show our findings in the context of tumor evolution, we use our 3D model to imitate tumor growth. We consider 3000 cells of (A) and (B) randomly positioned in the center of the 3D medium and let them grow according to the aforementioned rules. For two similar populations, both of them will appear at the surface and probable fixation is unlikely to happen due to the geometry of the tumor ( see FIG 3(a)). Then we consider the non-identical scenario with R_B=1.1 R_A. Based on the neutral theory, (A) should actively participate in the growth and should be present at the border of the tumor, in spite of having lower fitness. Based on Darwinian evolution, (B) is dominant and it will take over the whole tumor and will be the only population active in the border.
For the Eden model, as FIG 3(b) shows, (A) has a chance to appear at the border but finally it will be captured inside the tumor as the tumor becomes larger. When we add environmental barrier, the situation for (A) becomes worse and it will be run out of border faster (FIG 3(c)). However, when we add short-range migration by keeping N=10, the situation becomes completely different. First, two populations, as FIG 3(d) shows, became well-mixed. Then, in spite of being mixed, natural selection is not effective as before and (A) remains active in the border and participates in tumor evolution. Interestingly, while the dynamics of sub-clones has changed, the morphology of the tumor remains almost the same.
Figure 3: Evolutionary Dynamics of Tumor in presence of short-range migration
 Our results provide evidence for the possibility of violation of natural selection in well-mixed populations. These findings can be verified, thank recent experimental advancements.
Our results provide evidence for the possibility of violation of natural selection in well-mixed populations. These findings can be verified, thank recent experimental advancements.
© 2026 - The Mathematical Oncology Blog