Games and the Treatment Convexity of Cancer
Behind the Paper
Games and the Treatment Convexity of Cancer
Péter Bayer & Jeffrey West
Read the paper Open-access preprintTreatment convexity
Dose response curves are rarely linear: they typically are convex (figure 1B), concave (figure 1C) or sigmoidal shaped. By Jensen’s inequality, if the dose response function is convex, then even dosing is most effective (figure 1, purple). In contrast, if dose response function is concave, then uneven dosing is most effective. This simple observation relates to Nassim Taleb's concept called antifragility, which aims to map the relationship between input variance/volatility and output variance/volatility.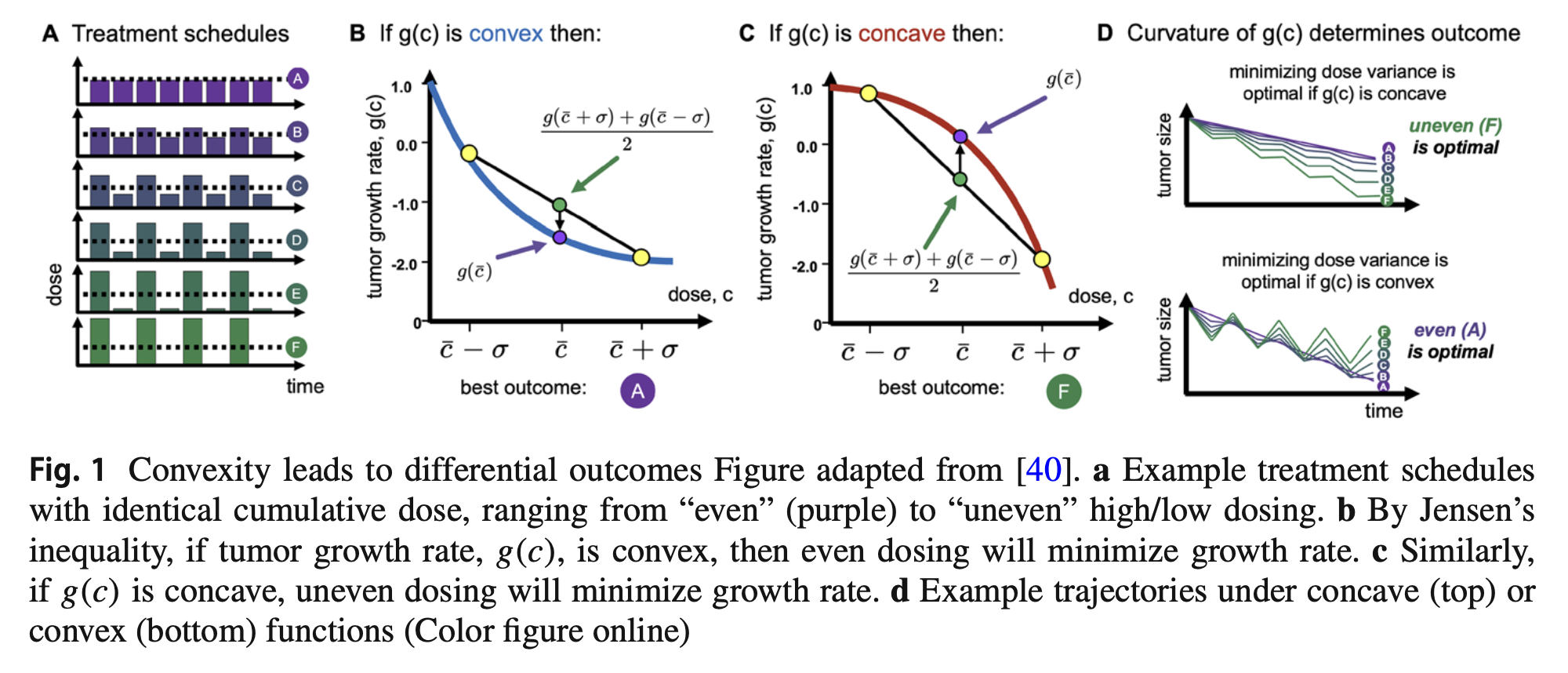
Ecological antifragility
In modeling cancer treatment response, we introduced the term "ecological antifragility" to refer to the mapping between dosing volatility and curvature [2], because this mapping considers the drug response of a single cell type in isolation and ignores heterogeneity and cancer evolution. Ecological antifragility provides a useful starting point when designing therapeutic regimens for single and combination treatments. In addition to response curvature, there is another way to think about ecological antifragility. In a paper published by Taleb in 2018, he describes antifragility in terms of threshold effects (see Figure 2) [3]. If some desired threshold is required, then only the right-tail of outcomes exceed the threshold. In fact, if the distribution variance is so low, none of the outcomes will exceed the threshold. In all cases, the mean is identical, indicating the utility of considering both the mean and variance. This applies straightforwardly to cancer care. For example, there is a desired threshold of tumor eradication, below which we achieve cure. It may be beneficial to widen the distribution of outcomes in order to increase the number of patients who exceed this threshold effect. Of course, this may come at the risk of also maximizing likelihood of the worst-of-the-worst poor outcomes, too, so one must take care. Back to game theory: we hypothesized that threshold effects may come into play in game theory, where a certain threshold of dosing may lead to a qualitatively change in game class. In contrast to ecological antifragility, evolutionary antifragility explicitly considers competition between heterogeneous subtypes within a tumor. This evolutionary antifragility quantifies the effect of cell-cell interactions on second-order effects of dosing.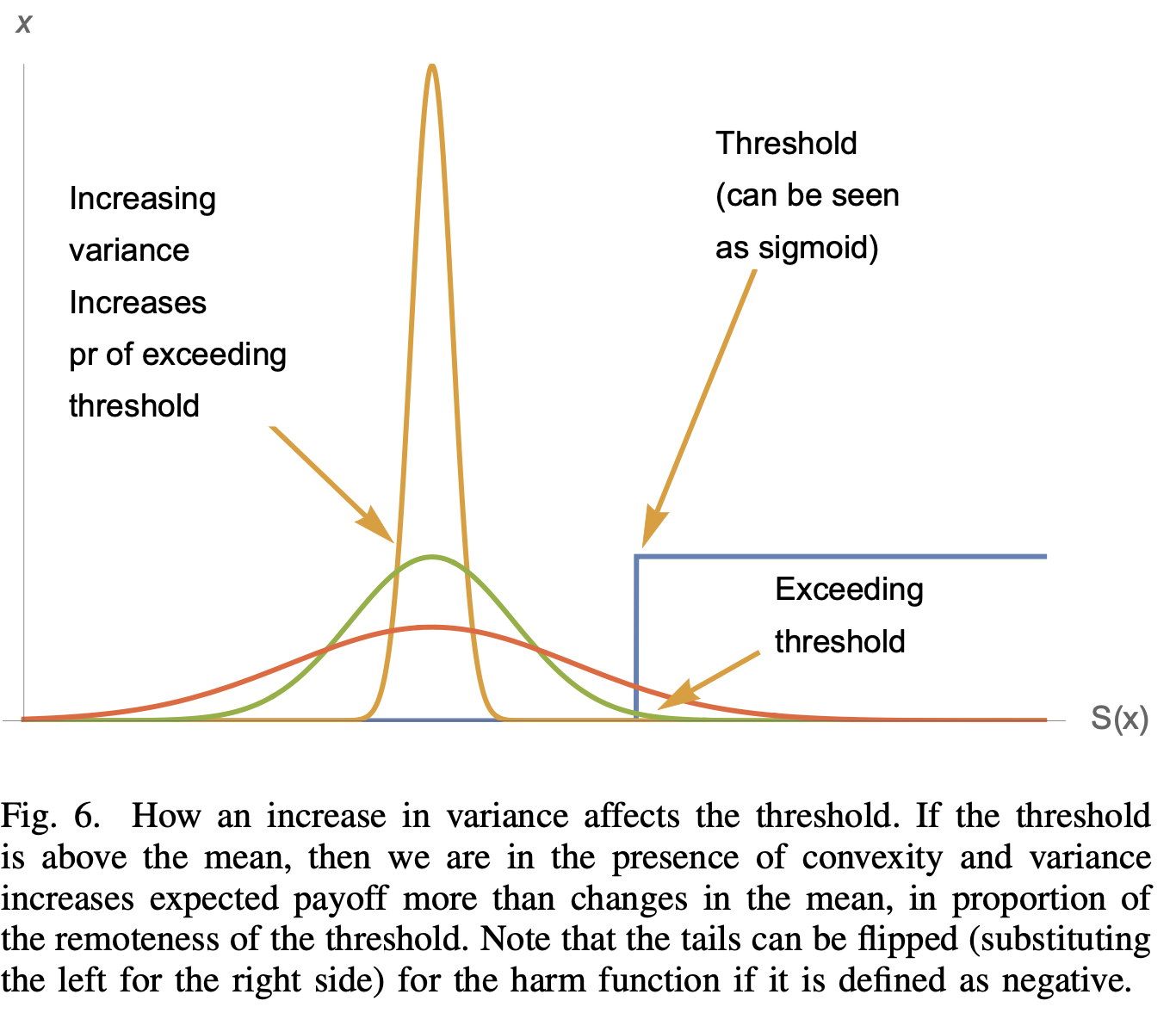
Evolutionary antifragility
We construct an evolutionary game theory model describing the competition/cooperation between two competing cell types: type 1 and 2. We start off by defining two parameters: Δ1, denoting the payoff advantage of playing 1 instead of 2 against an opponent with strategy 1, and Δ2, the payoff advantage of playing 2 instead of 1 against an opponent with strategy 2. This allows us to visualize the game space on two axes: Δ1 and Δ2, whereby four quadrants exist: 1) Prisoner’s Dilemma (type 2 is the growth-optimal strategy), 2) Coordination (type 1 and 2 have bi-stability), 3) Anti-Coordination (stable coexistence of the two cell types) and 4) Harmony (type 1 is the growth-optimal strategy). Next, we consider the effect of treatment on the competitive dynamics between the two types. In general, games are understudied in cancer, and typically only measured quantifiably at sparse dose values. While the direct relationship between dose and game space is unknown, we made a parsimonious assumption that treatment represents a linear projection in game space (see Figure 2A, below). This assumption not without basis, as one previous study (reproduced in Figure 2B) appears to show a linear relationship between dose and game space (see Figure 7B in the Farrokhian preprint [4]) under etoposide dosing (now published in Science Advances [5]).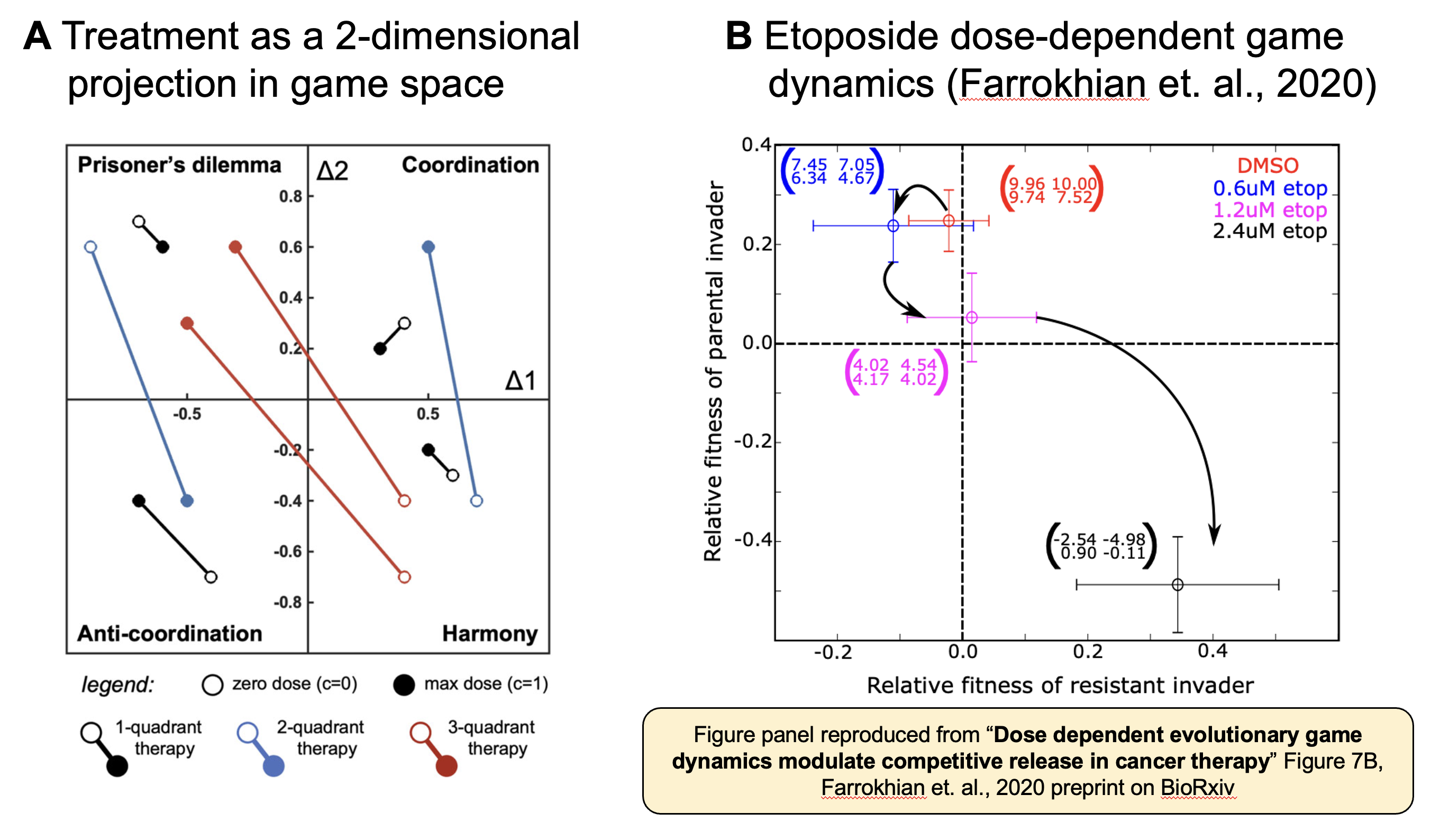
Interventional antifragility
While, we don't go into it in this manuscript, the idea to combine evolutionary game theory and treatment convexity were inspired by the concept of adaptive therapy. Here's the logic:- Adaptive therapy trials were designed to capitalize on a cost of resistance by treating less often, in order to maintain a stable population of treatment-sensitive cells
- Evolutionary game theory is commonly used to define the nature of resistance cost and/or cell-cell interactions in cancer
- Adaptive therapy minimizes both the cumulative dose (a first-order effect) and increases the dose variance (a second-order effect)
- Therefore, it's important to study second-order effects using evolutionary game theory models
References
- Bayer, P., West, J. Games and the Treatment Convexity of Cancer. Dyn Games Appl (2023). https://doi.org/10.1007/s13235-023-00520-z.
- West, Jeffrey, et al. "Antifragile therapy." BioRxiv (2020): 2020-10. https://doi.org/10.1101/2020.10.08.331678.
- Taleb, Nassim Nicholas. "(anti) fragility and convex responses in medicine." Unifying Themes in Complex Systems IX: Proceedings of the Ninth International Conference on Complex Systems 9. Springer International Publishing, 2018.
- Farrokhian, N., et al. "Dose dependent evolutionary game dynamics modulate competitive release in cancer therapy." bioRxiv 2020.18.303966 (2020).
- Farrokhian, Nathan, et al. "Measuring competitive exclusion in non–small cell lung cancer." Science Advances 8.26 (2022): eabm7212.
© 2026 - The Mathematical Oncology Blog