Space Accelerates Evolution
Realized Darwinian fitness
Tissue structure accelerates evolution: premalignant sweeps precede neutral expansion
Jeffrey West, Ryan O. Schenck, Chandler Gatenbee, Mark Robertson-Tessi, Alexander R. A. Anderson
Read the manuscript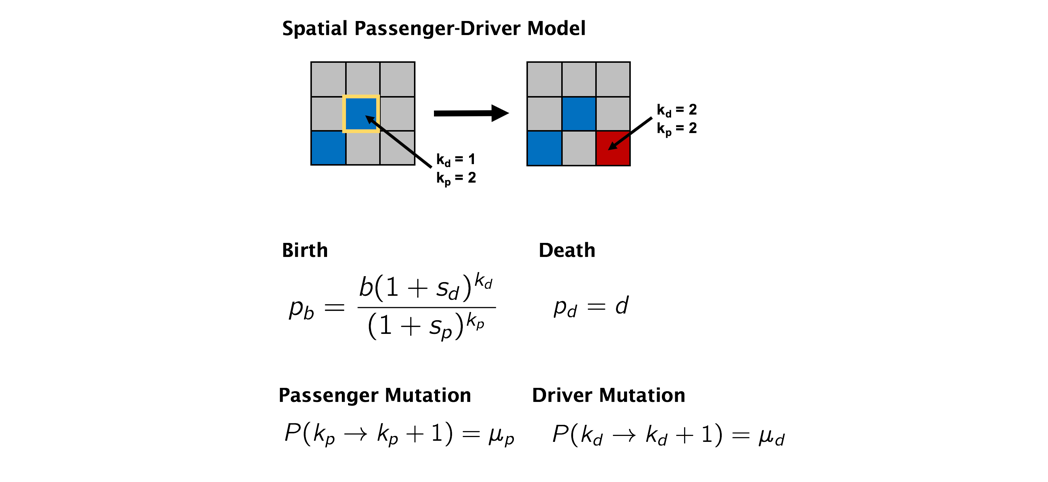
Constraining spatial domain sizes
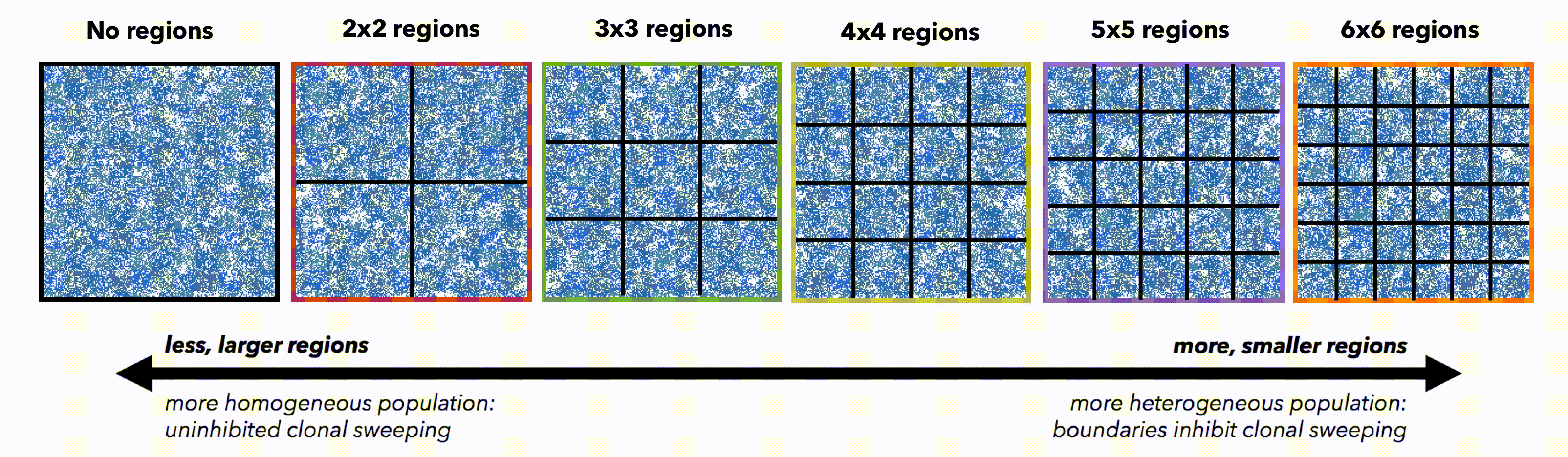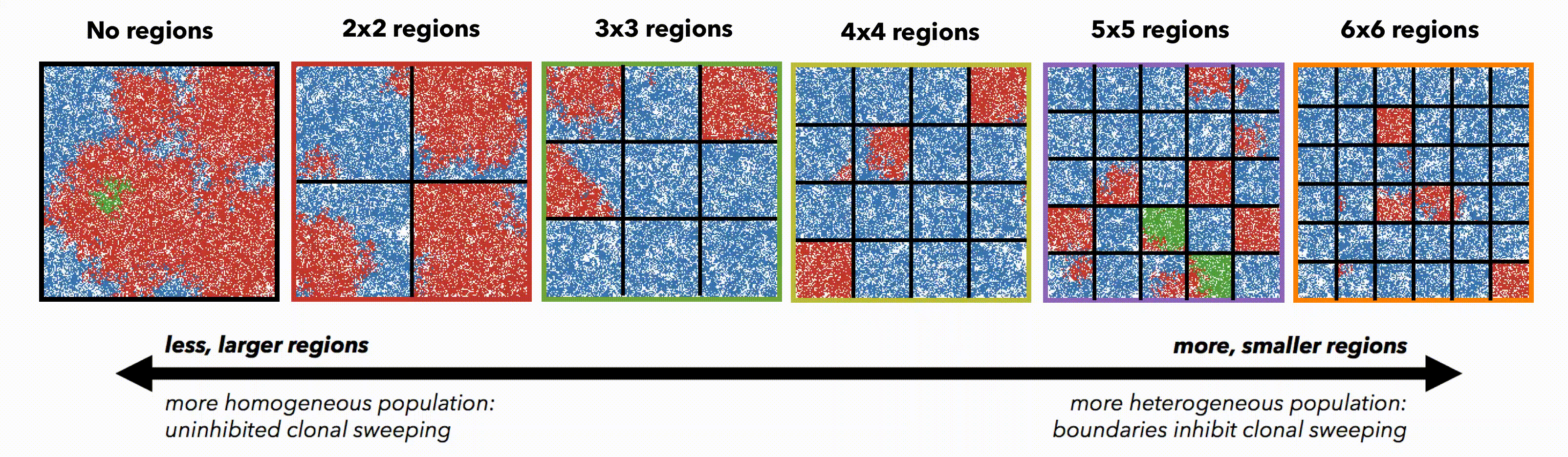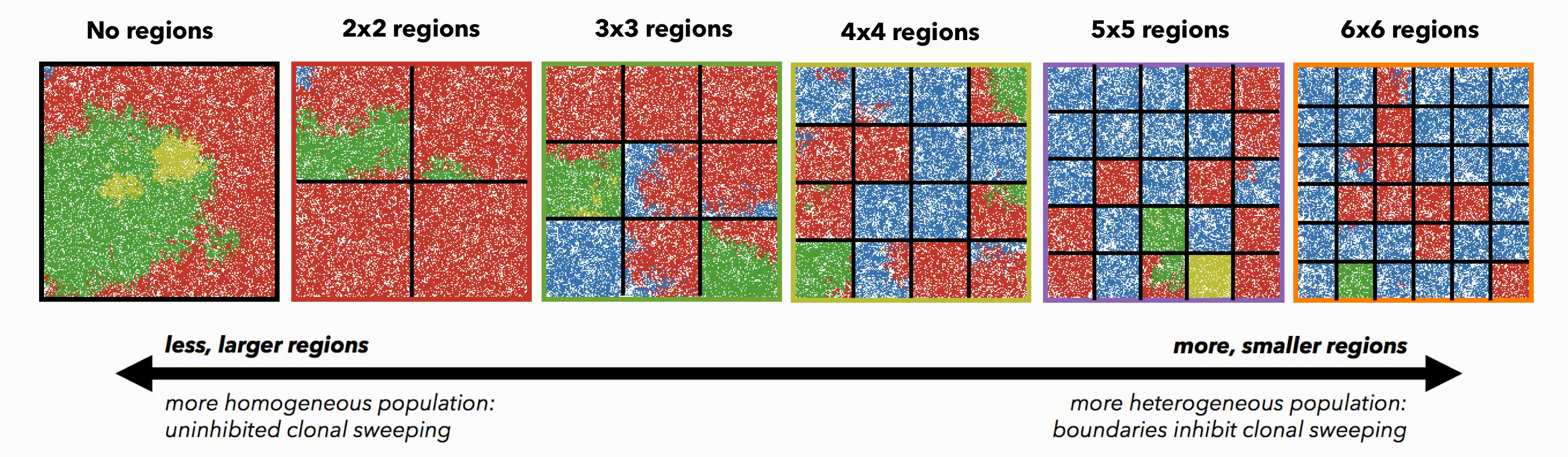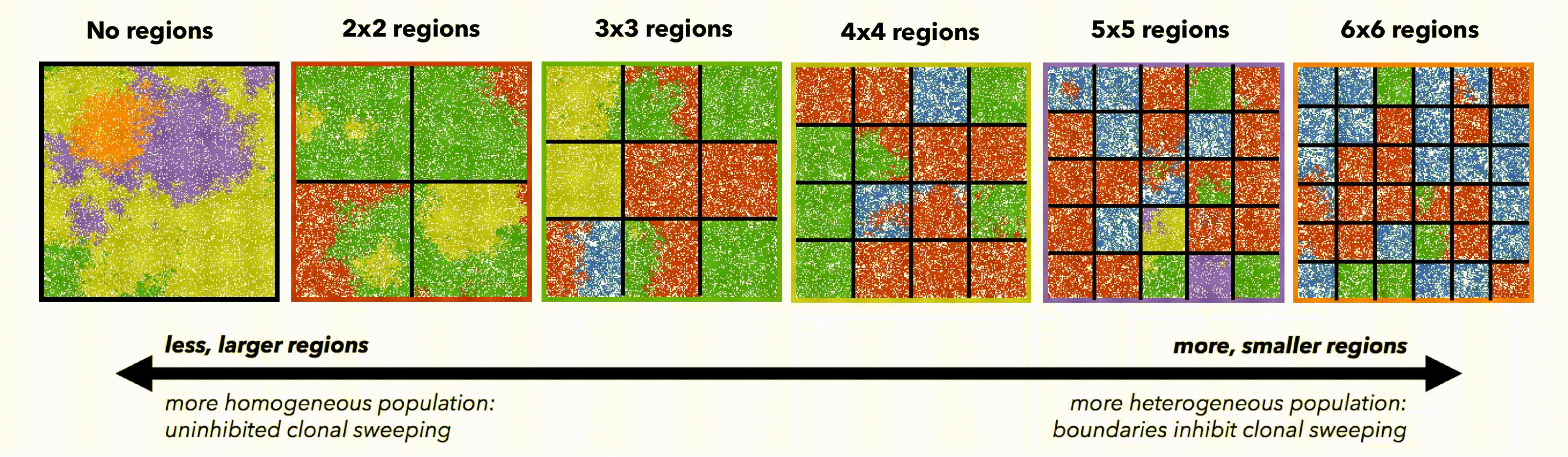
Tumors constrained to smaller domain sizes (figure 2, left) show consistently lower driver and passenger diversity than for larger domain sizes. Small, tightly-coupled homogeneous populations of cells are able to quickly sweep each successive driver mutation. Larger domains consist of a heterogeneous population; with many more cell divisions, the odds of accruing another driver mutation are increased, but they have little chance of sweeping through the large domain. Fig.2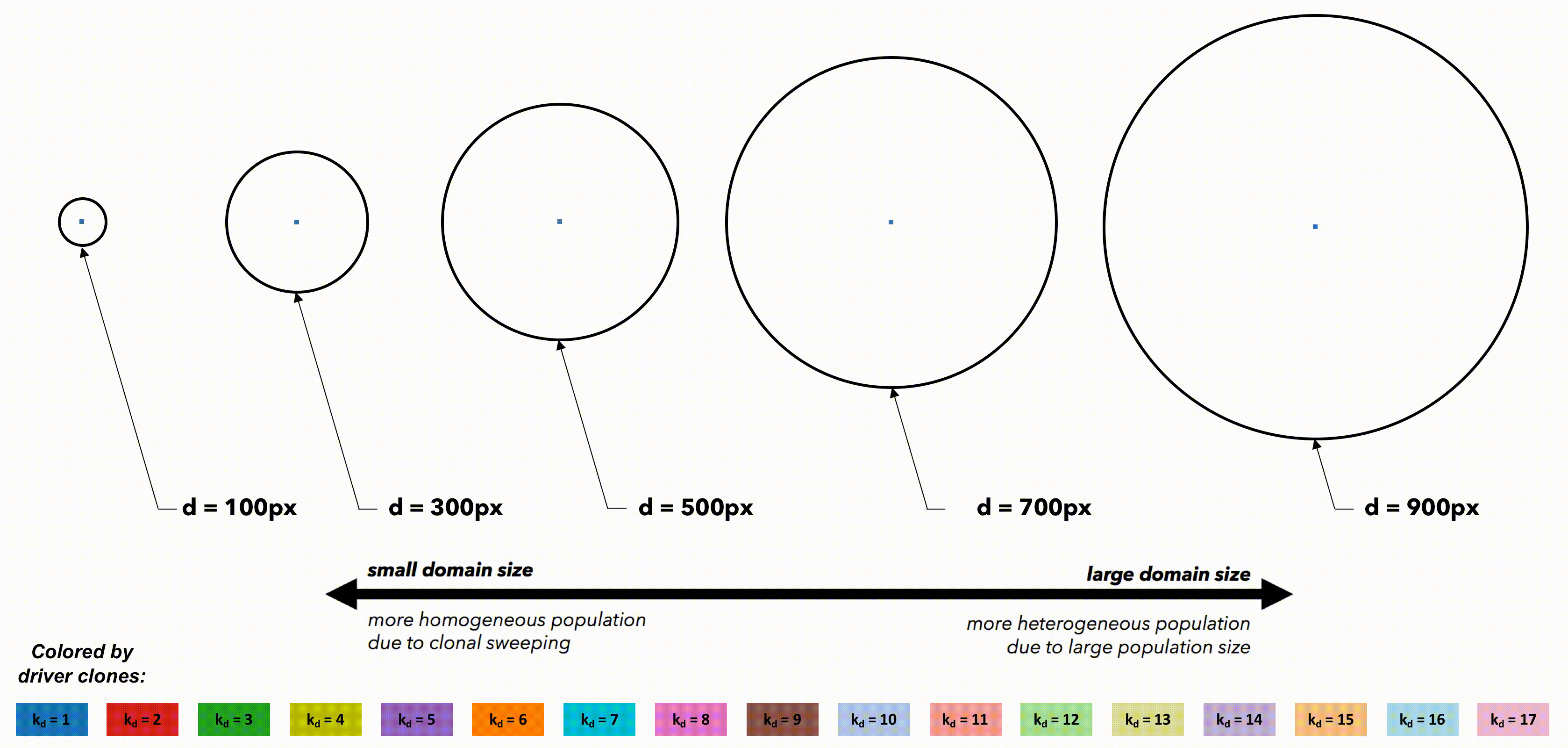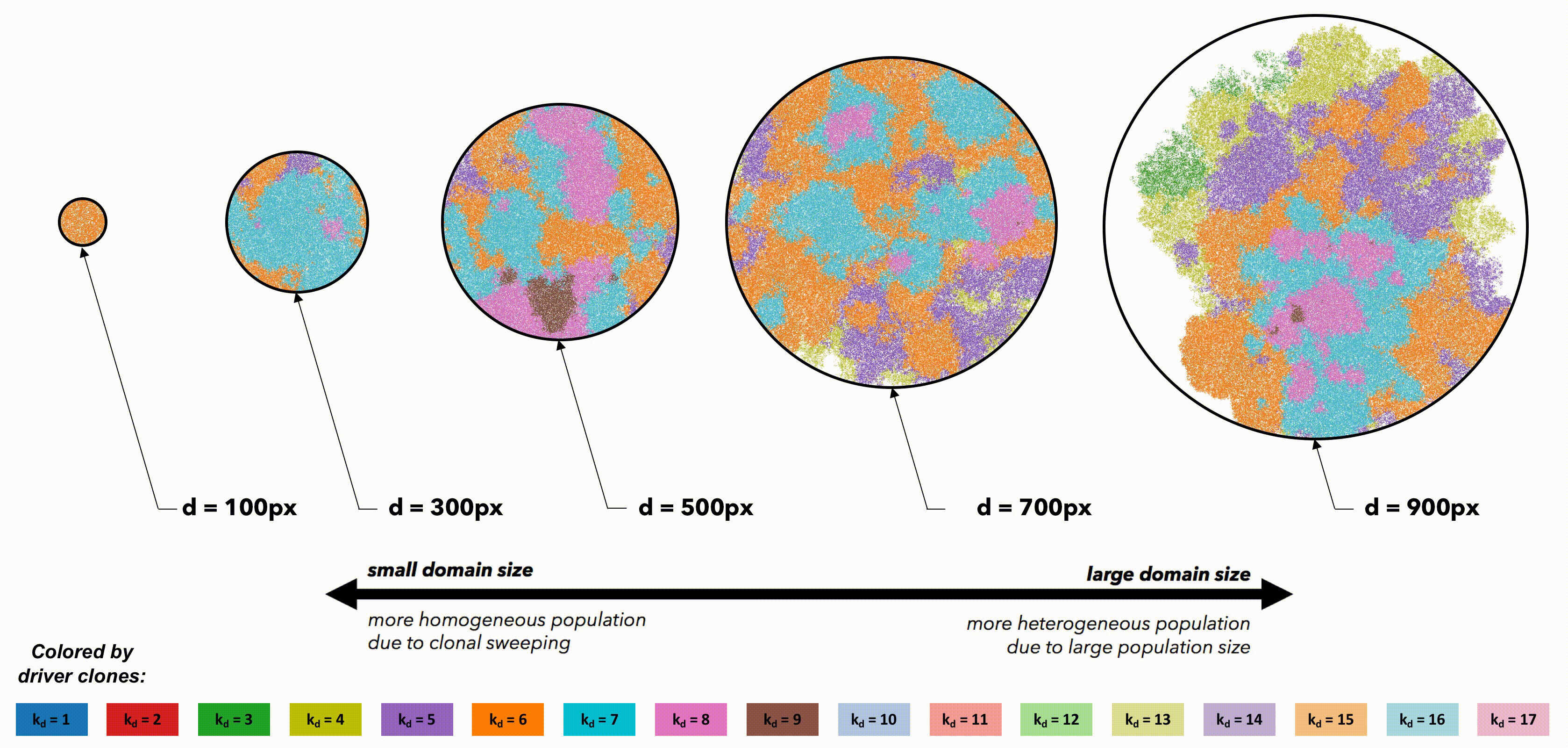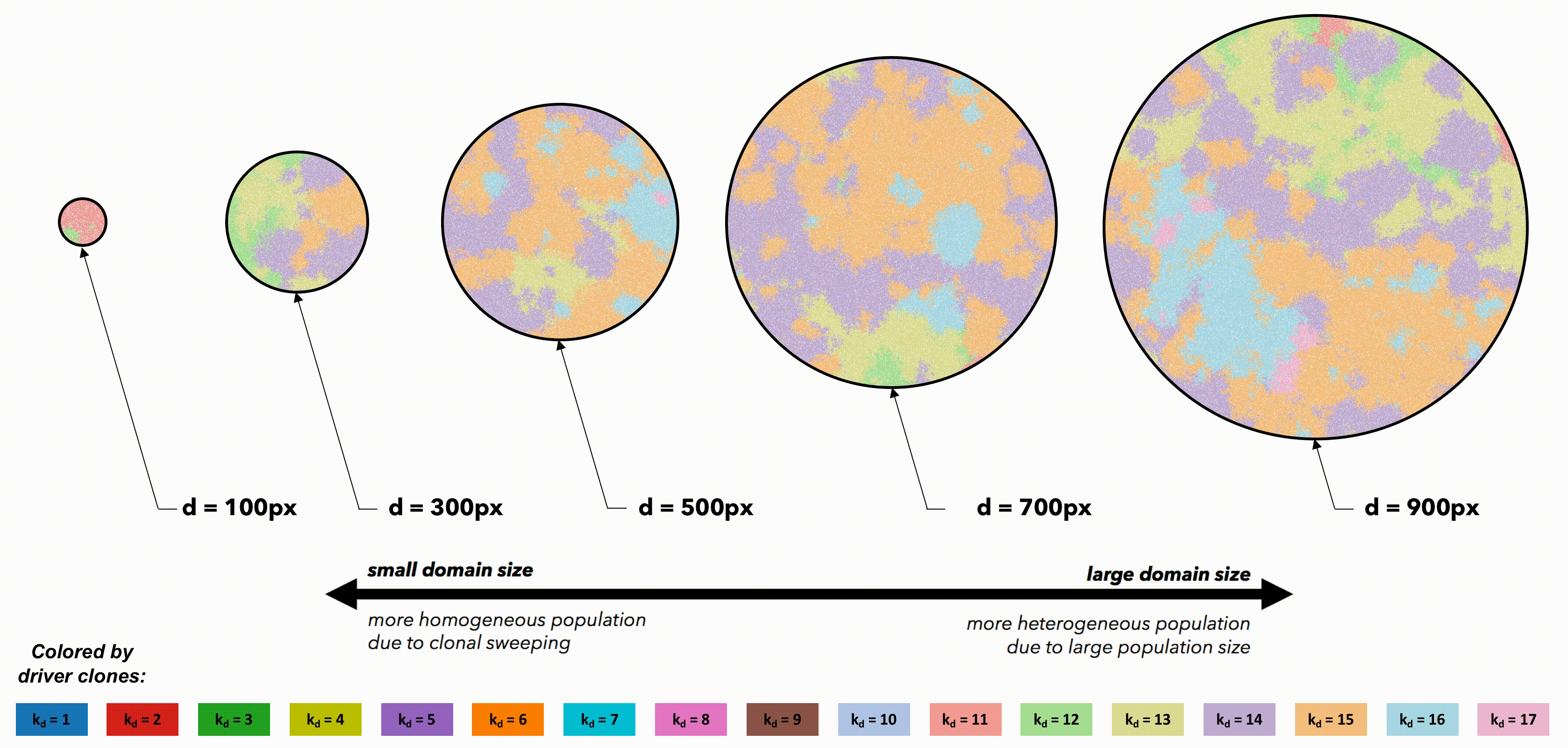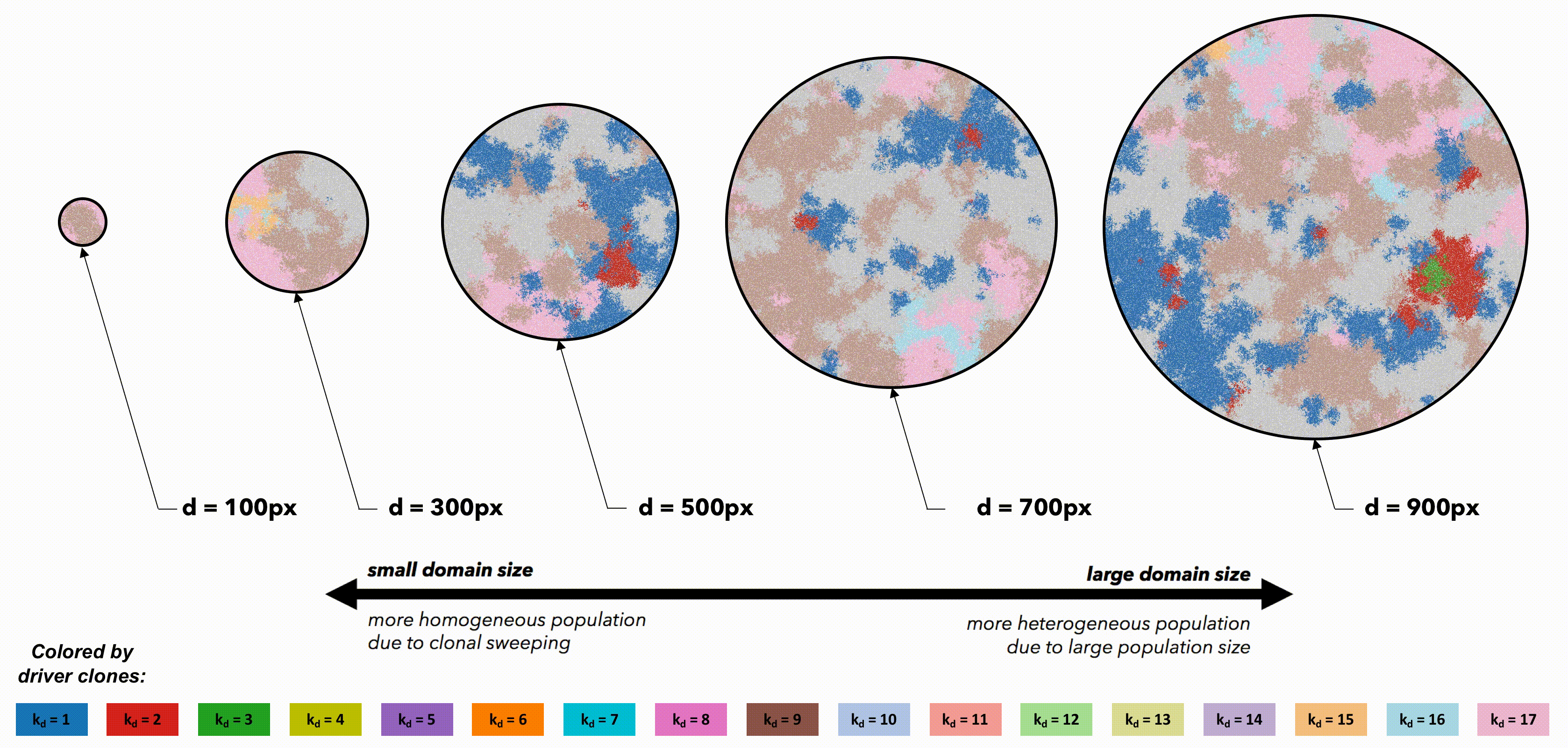 But perhaps these differences are simply due to population size. To control for population size effects in this evolutionary arms race, identical domains are segregated into non-interacting regions of varying size (figure 3). Again, smaller (highly constrained) regions provide a stronger selection force, sweeping away weaker subclones. This sweeping stops at each region's boundaries, resulting in a heterogeneous population of locally homogeneous regions. Boundaries limit new clones from expanding beyond a single region, decreasing the average number of drivers in the population.
Fig.3
But perhaps these differences are simply due to population size. To control for population size effects in this evolutionary arms race, identical domains are segregated into non-interacting regions of varying size (figure 3). Again, smaller (highly constrained) regions provide a stronger selection force, sweeping away weaker subclones. This sweeping stops at each region's boundaries, resulting in a heterogeneous population of locally homogeneous regions. Boundaries limit new clones from expanding beyond a single region, decreasing the average number of drivers in the population.
Fig.3
 Bounded, non-interacting regions play a role in human cancers, which are often locally constrained to a single gland or a duct. Such glandular or ductal structures allow for limited cellular mixing during premalignant growth, enabling the tumor to explore new (and often less constrained) environments. In figure 4 each segregated region may now circulate cells into a neighboring region at a low or high rate of mixing (left and right columns, respectively).
Fig.4
Bounded, non-interacting regions play a role in human cancers, which are often locally constrained to a single gland or a duct. Such glandular or ductal structures allow for limited cellular mixing during premalignant growth, enabling the tumor to explore new (and often less constrained) environments. In figure 4 each segregated region may now circulate cells into a neighboring region at a low or high rate of mixing (left and right columns, respectively).
Fig.4
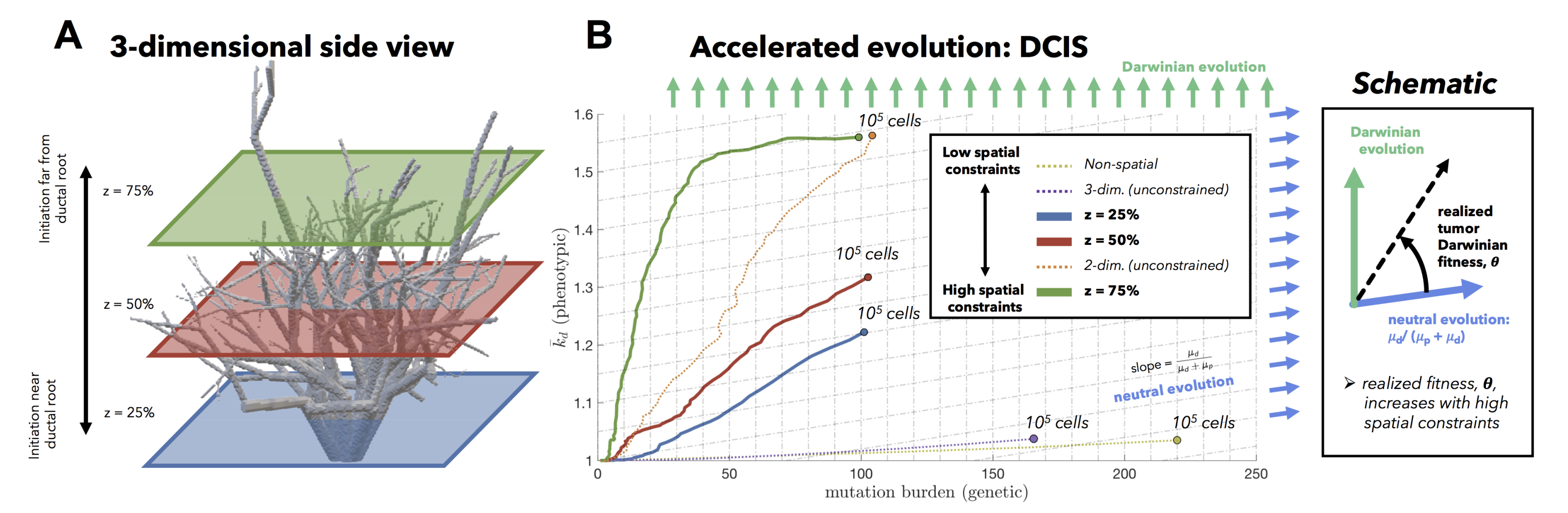
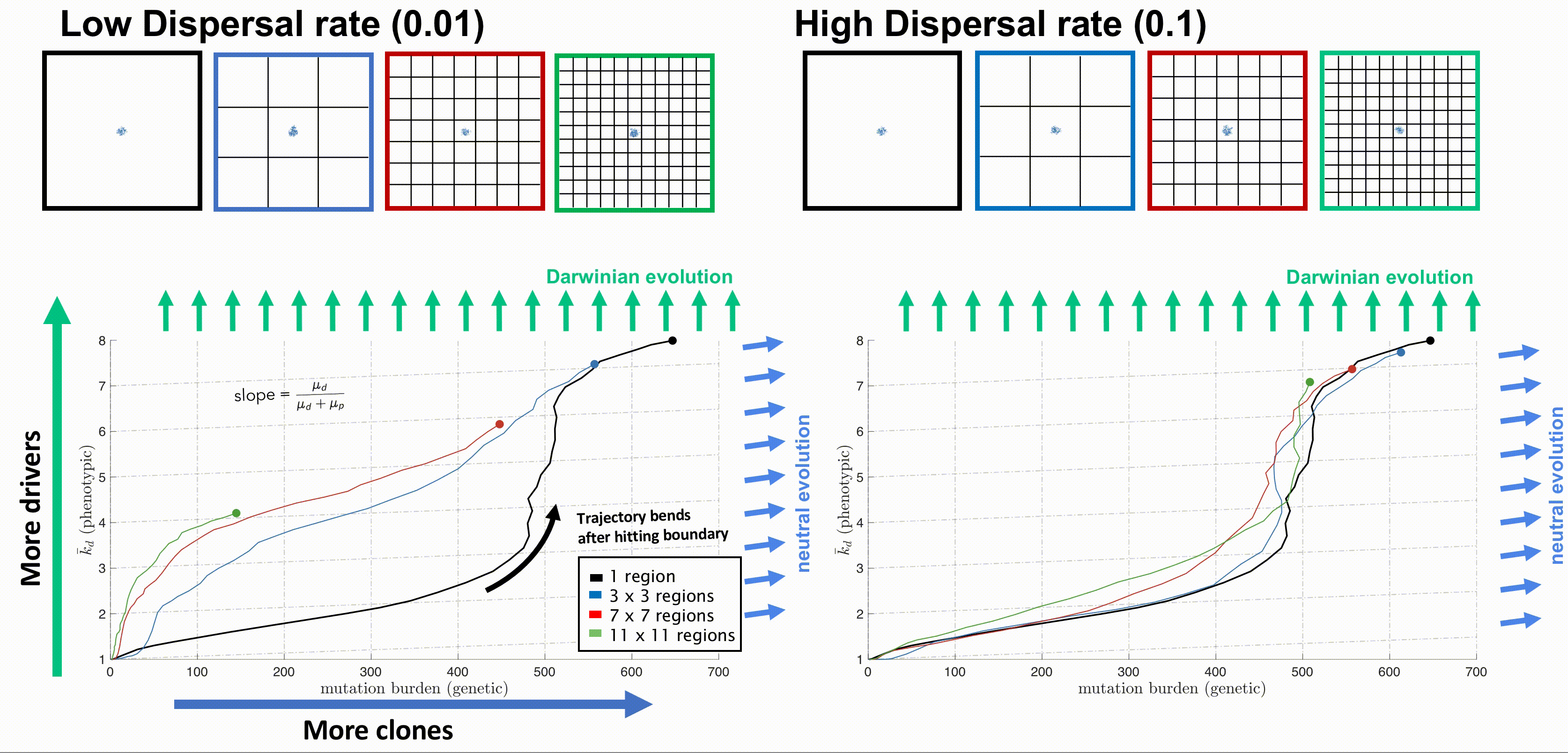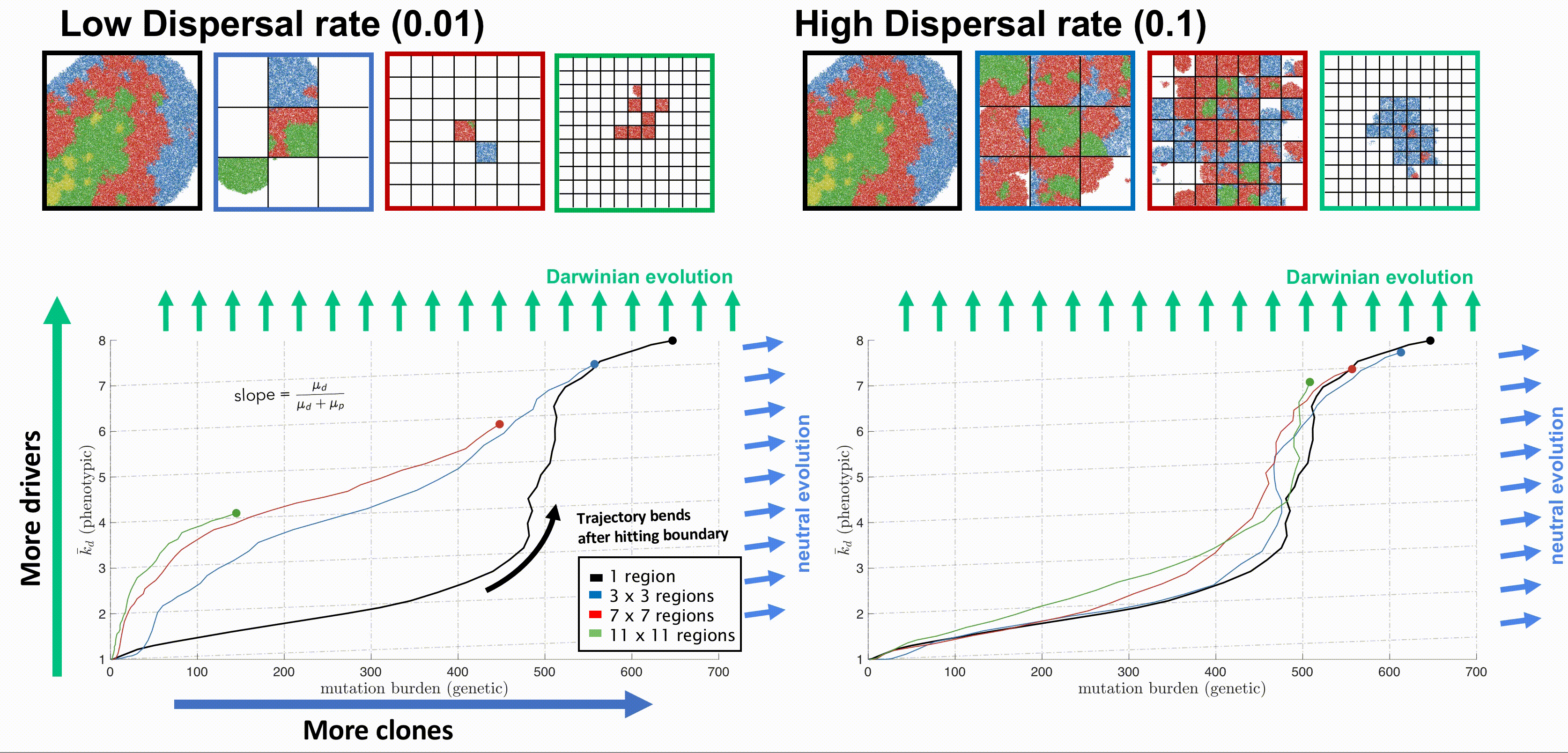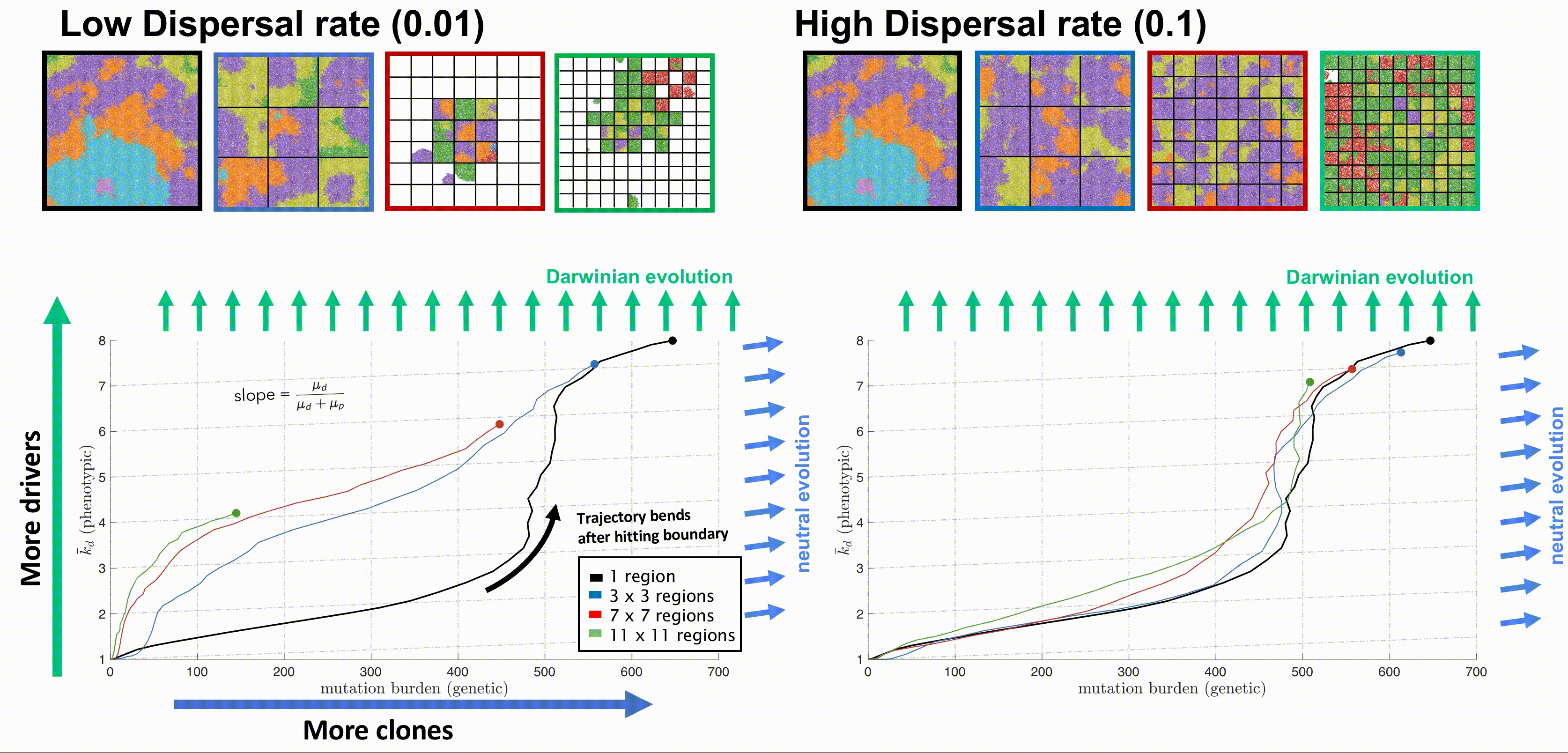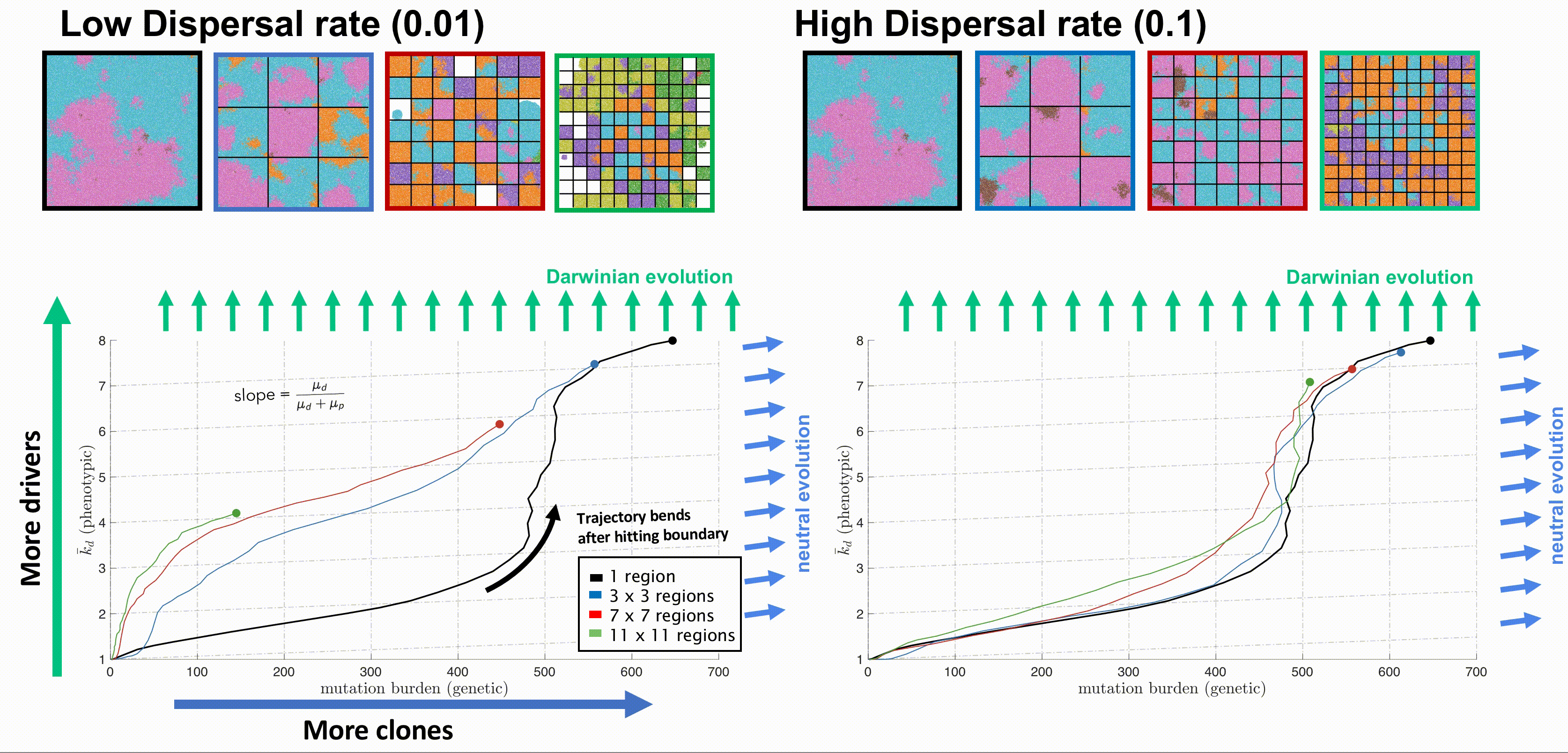 Tumors evolve on two axes: genetic (mutation burden; x-axis) and phenotypic (average number of driver mutations, \(\bar{k}_d\); y-axis). The evolution of an unsegregated tumor in is shown in black, accumulating genetic diversity (left-to-right) over time with a slow accumulation of drivers (bottom-to-top). This state space diagram allows us to track the accelerated acquisition of drivers in a precancerous population, with respect to tumor size. The trajectory of a tumor's evolution quantifies the tumor-scale effect of domain size and mixing, over time. Despite seeding simulations with identical parameterization, tumors may evolve in a neutral or Darwinian mode. Neutral tumors acquire drivers at a rate equal to the ratio of drivers to all mutations (blue arrows). Conversely, Darwinian tumors sweep each new driver mutation through the population, resulting in a vertical trajectory (green arrows).
Tumors evolve on two axes: genetic (mutation burden; x-axis) and phenotypic (average number of driver mutations, \(\bar{k}_d\); y-axis). The evolution of an unsegregated tumor in is shown in black, accumulating genetic diversity (left-to-right) over time with a slow accumulation of drivers (bottom-to-top). This state space diagram allows us to track the accelerated acquisition of drivers in a precancerous population, with respect to tumor size. The trajectory of a tumor's evolution quantifies the tumor-scale effect of domain size and mixing, over time. Despite seeding simulations with identical parameterization, tumors may evolve in a neutral or Darwinian mode. Neutral tumors acquire drivers at a rate equal to the ratio of drivers to all mutations (blue arrows). Conversely, Darwinian tumors sweep each new driver mutation through the population, resulting in a vertical trajectory (green arrows).
Modeling the acceleration of evolution by tissue structure
We now extend this generalized understanding of the model to a more biologically realistic setting: the 3-dimensional branching topology of a breast ductal network spatial structure representing biologically realistic timescales (~500 to 1500 days), biologically realistic cell numbers for precancerous lesions (100,000 cells), and biologically realistic spatial topologies (ductal network structures). Depending on the initial tumor size (say, the star in figure 5), the tumor is subject to topology-dependent cellular mixing and spatial constraints. We can run the model again, constrained to the topology of the ductal network (figure 6).
Fig.5

Fig.6
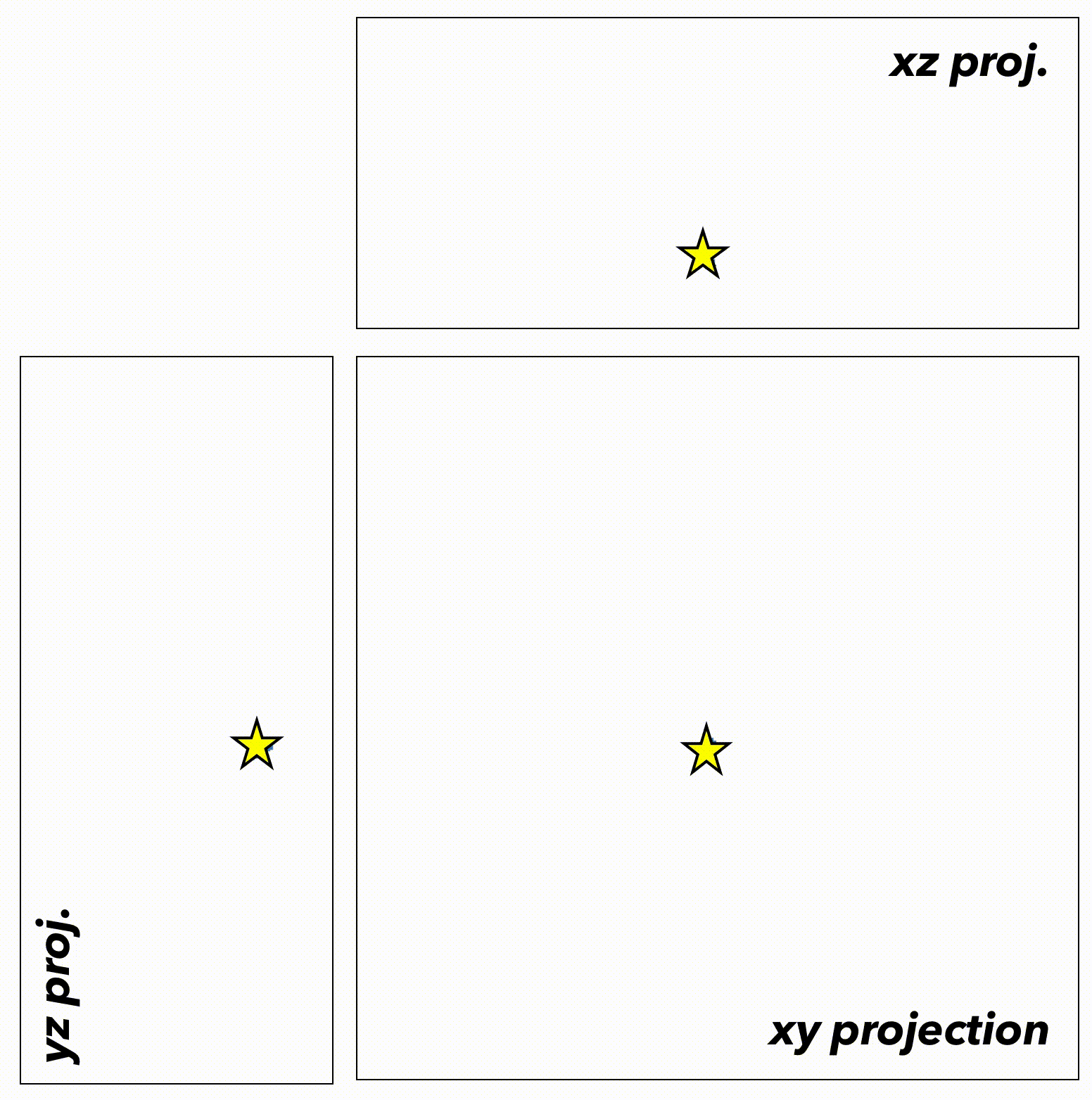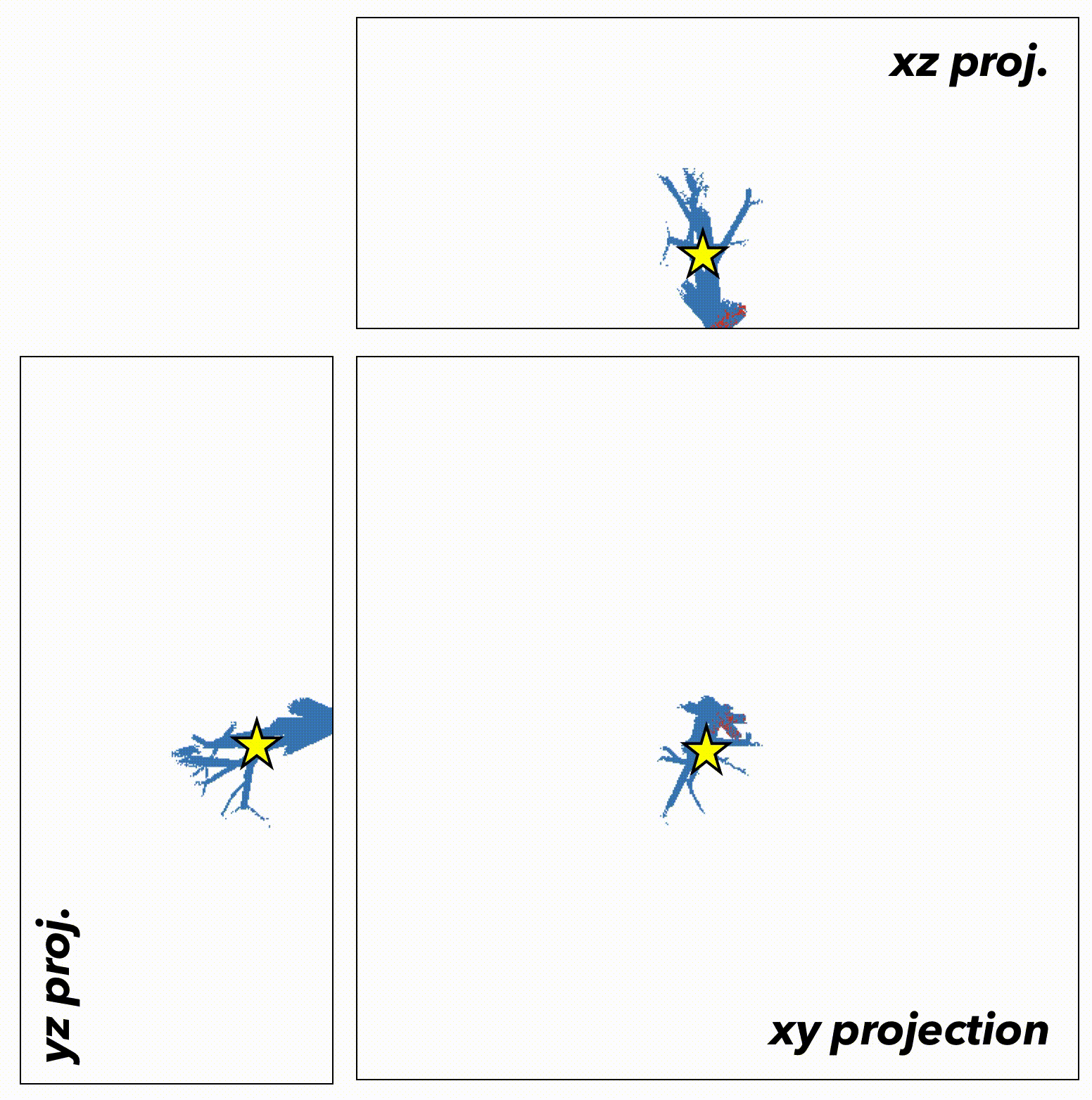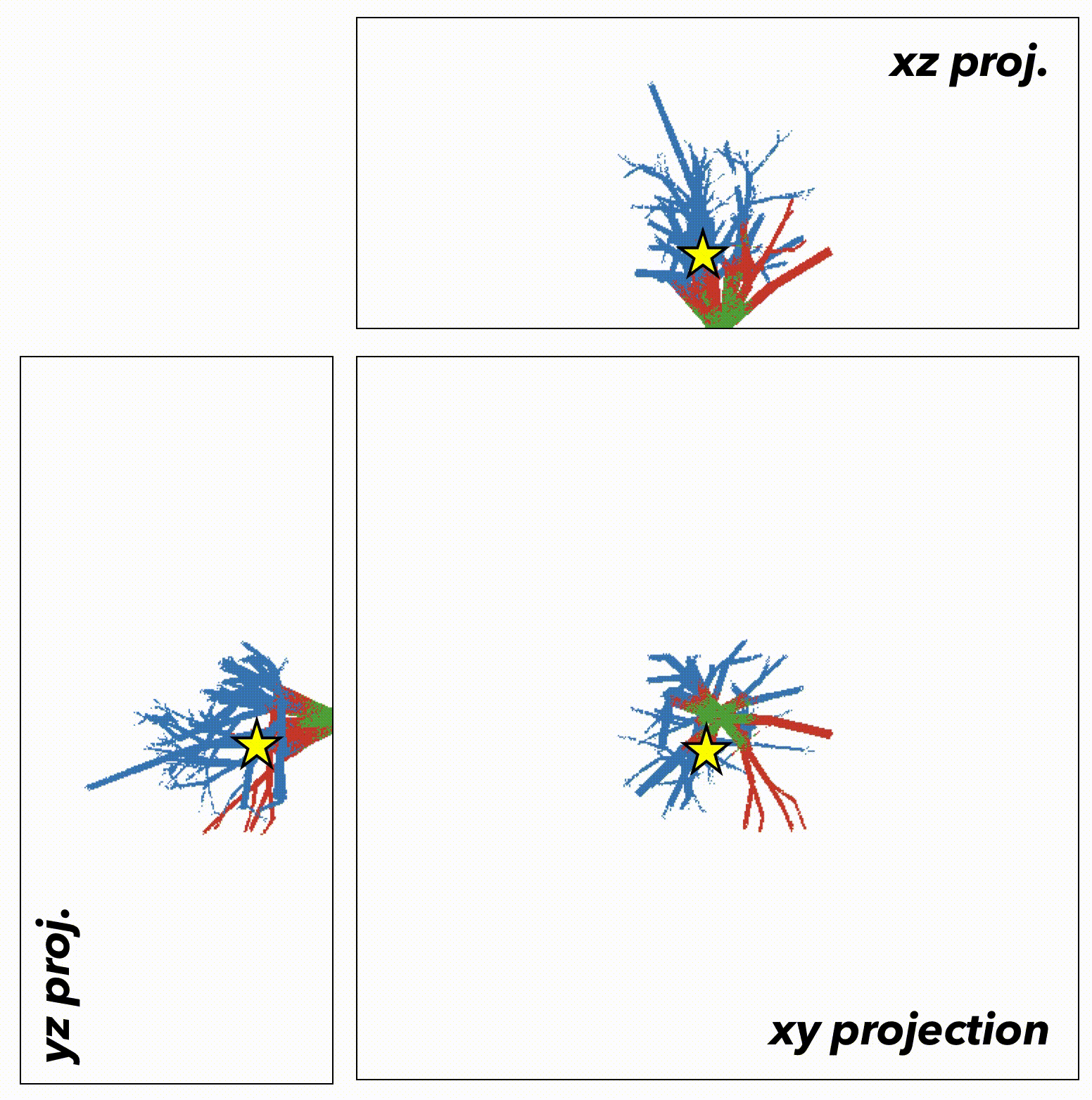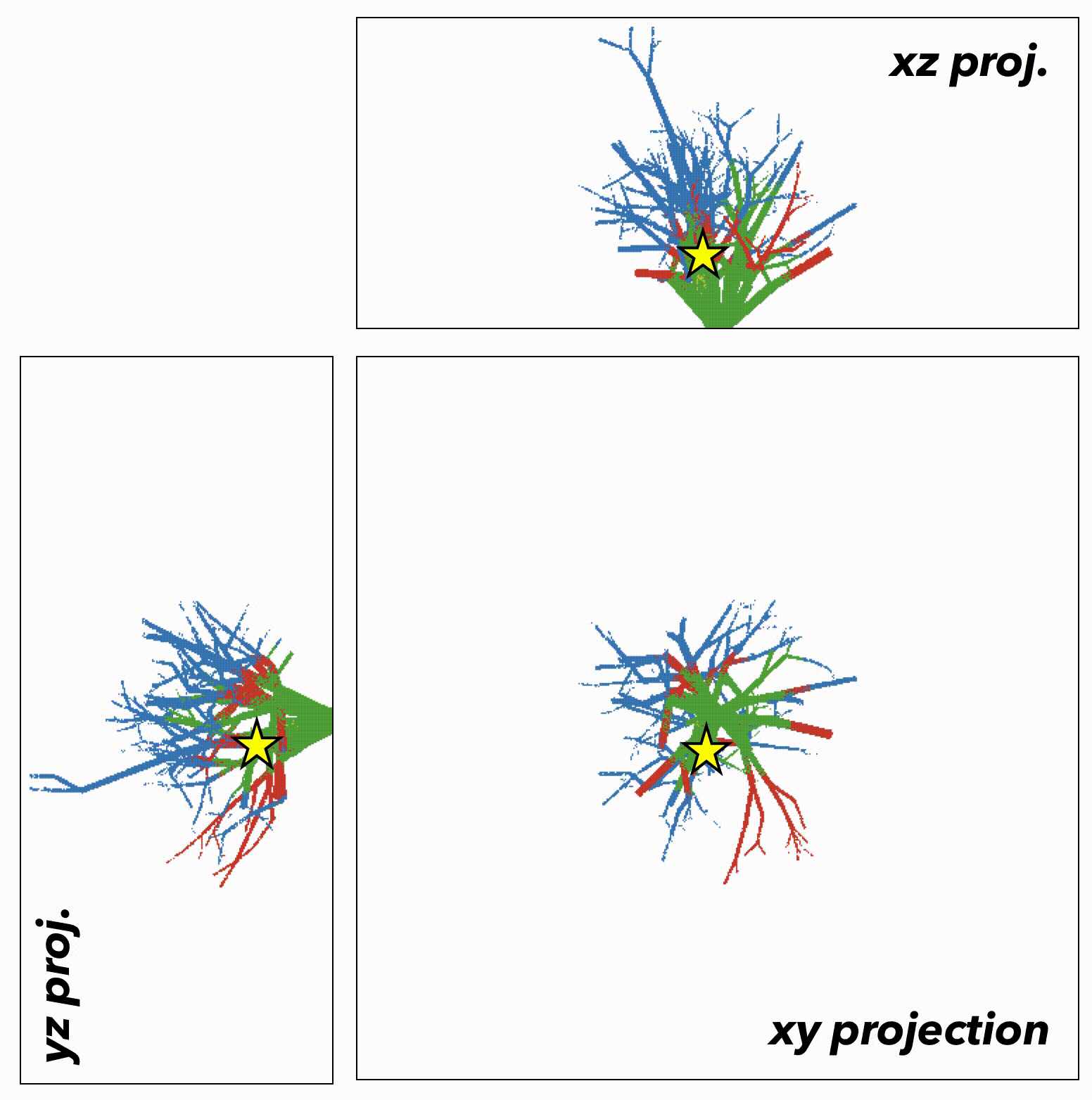
 In this case, the important result may have been lost without spatially-explicit models. In a non-spatial setting, fitter clones may rarely be constrained from expanding, with significant genetic hitch-hikers. Contrasting that to a spatial setting where many fitter clones may indeed exist – but are unable to sweep. In some cases, space may actually help the sweeping by enabling a Goldilocks zone of cellular mixing and negative selection allowing expansion and sweeping to occur simultaneously.
In this case, the important result may have been lost without spatially-explicit models. In a non-spatial setting, fitter clones may rarely be constrained from expanding, with significant genetic hitch-hikers. Contrasting that to a spatial setting where many fitter clones may indeed exist – but are unable to sweep. In some cases, space may actually help the sweeping by enabling a Goldilocks zone of cellular mixing and negative selection allowing expansion and sweeping to occur simultaneously.
References
- Tissue structure accelerates evolution: premalignant sweeps precede neutral expansion. Jeffrey West, Ryan O. Schenck, Chandler Gatenbee, Mark Robertson-Tessi, Alexander R. A. Anderson bioRxiv 542019; doi: https://doi.org/10.1101/542019
© 2026 - The Mathematical Oncology Blog