Balanced phenotypic trade-offs in polyploid cancer populations slow down invasion
Behind the paper
Integrating mathematical modeling with high throughput imaging explains how polyploid populations behave in nutrient-sparse environments
Gregory J. Kimmel, Mark Dane, Laura M. Heiser, Philipp M. Altrock and Noemi Andor
Read the paper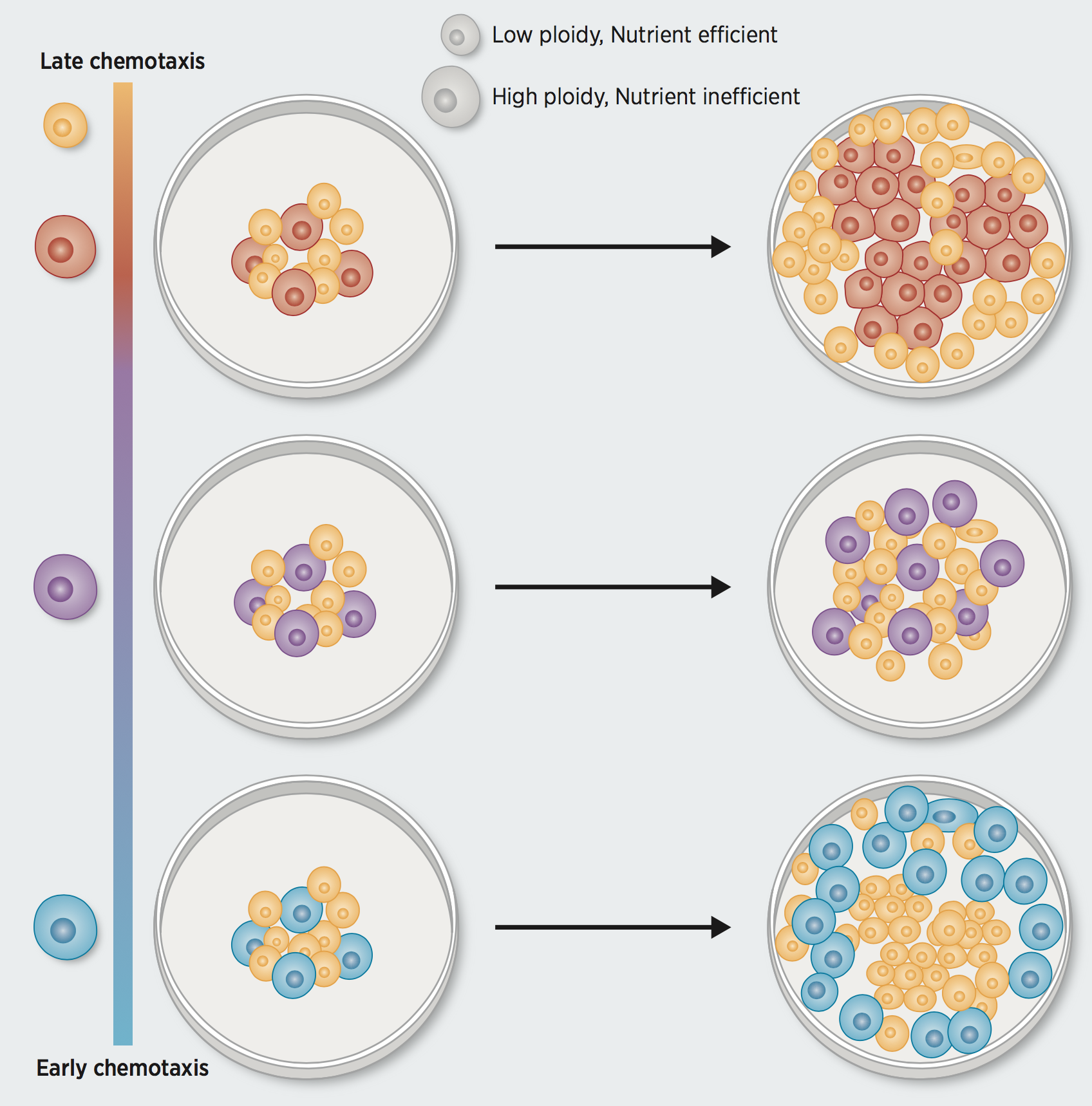
Figure 1. Imbalance between nutrient efficacy and chemotactic superiority accelerates invasion of polyploid populations: subpopulation will segregate and only one type dominates at the invasion front.
When calibrating our model to a given ECM environment, growth patterns of a substantial fraction of replicates of that ECM could not be explained by fixed choices of energy sensitivity. A potential explanation for this is variable cell compositions across experimental replicates. The observation of a bimodal distribution in the DNA content of S-phase cells strengthened the hypothesis that HCC1954 is a polyploid cell line. One event that could have led to clones of variable ploidies coexisting in this cell line is WGD. Selection pressures for WGD suggests that it mitigates the accumulation of deleterious somatic alterations (2). However, there may be an energetic cost the cells pay for this robustness. In line with this, we find that high-ploidy breast cancer cell lines are resistant to cytotoxic drugs, but tend to be more sensitive to inhibitors of mTOR, EGFR, and MAPK signaling pathways. We used our model to investigate how variable energetic drawbacks of additional DNA content, affects competitiveness of high-ploidy cells as members of a polyploid population. We find that long-term coexistence of low- and high-ploidy clones occurs when sensitivity of the latter to energy scarcity is well-offset by their chemotactic ability to populate new terrain (3). Only when the high-ploidy clone has an intermediate motility, does it persistently coexist with the low-ploidy clone, both at the center and edge of the dish. Delaying chemotactic response of highly chemotactic cells could slow down invasion by maximizing competition within a polyploid population. If, on the other hand, chemotactic response of high-ploidy cells is already at an intermediate level, our simulation suggests that further reduction may accelerate invasion of low-ploidy cells. In contrast to cell lines, WGD events in primary tumors are mostly clonal, not subclonal. Clones carrying a doubled genome often sweep over the population, such that by the time the tumor is detected, the diploid ancestor no longer exists. A related scenario is advanced, therapy-exposed tumors shown to revert to genomic stability, potentially bringing a WGD population back to a genomic state that more closely resembles its diploid ancestral state (4). The model presented here can investigate how dynamics between the two subpopulations unfold in both of these scenarios—early, shortly after the WGD or late, after therapy exposure. This would characterize what circumstances prevent the WGD carrying clone from becoming dominant or from retaining its dominance and could help explain WGD incidence in primary and recurrent tumors.Further Reading
- Keenan AB, Jenkins SL, Jagodnik KM, Koplev S, He E, Torre D, et al. The Library of Integrated Network-Based Cellular Signatures NIH Program: System-Level Cataloging of Human Cells Response to Perturbations. Cell Syst. 2018;6:13–24.
- López S, Lim EL, Horswell S, Haase K, Huebner A, Dietzen M, et al. Interplay between whole-genome doubling and the accumulation of deleterious alterations in cancer evolution. Nat Genet. 2020;52:283–93.
- Kimmel GJ, Dane M, Heiser LM, Altrock PM, Andor N. Integrating mathematical modeling with high throughput imaging explains how polyploid populations behave in nutrient-sparse environments. Cancer Res [Internet]. American Association for Cancer Research; 2020 [cited 2020 Oct 5]; Available from: https://cancerres.aacrjournals.org/content/early/2020/09/16/0008-5472.CAN-20-1231
- Morrissy AS, Garzia L, Shih DJH, Zuyderduyn S, Huang X, Skowron P, et al. Divergent clonal selection dominates medulloblastoma at recurrence. Nature. 2016;529:351–7.
© 2026 - The Mathematical Oncology Blog